

El cenancestro se define como el último ancestro común de todos los seres vivos que existen en la actualidad. Su naturaleza se ha inferido a partir de la identificación de los genes homólogos entre los linajes de arqueas, bacterias y eucariontes. Estas inferencias indican que el cenancestro poseía un sistema de traducción de proteínas relativamente moderno y que era similar en complejidad a una célula actual. Sin embargo, las enzimas clave que se encargan tanto de la replicación del material genético como de la biosíntesis de las membranas celulares no son homólogas entre las bacterias, las arqueas y los eucariontes. Aquí revisamos brevemente la historia del concepto del último ancestro común y las distintas hipótesis que se han propuesto sobre su biología.
Los biólogos evolutivos topan con el mismo dilema lógico que los historiadores y sólo pueden presentar argumentos basados en la suposición de que, de todas las secuencias históricas plausibles, existe una que es más probable que sea una descripción correcta de los hechos del pasado.
Lynn Margulis, 1975, 29:21-38
El concepto de homología y el árbol universal de la vida
Dicen que una imagen vale más que mil palabras. En el caso de la teoría de la evolución, una de las imágenes más sugestivas que existen se encuentra en un cuaderno de notas escrito por Darwin tiempo antes de la publicación de El origen de las especies (figura 1). En este cuaderno se puede apreciar cómo Darwin representa mediante un dibujo el origen de distintas especies a partir de un ancestro común.
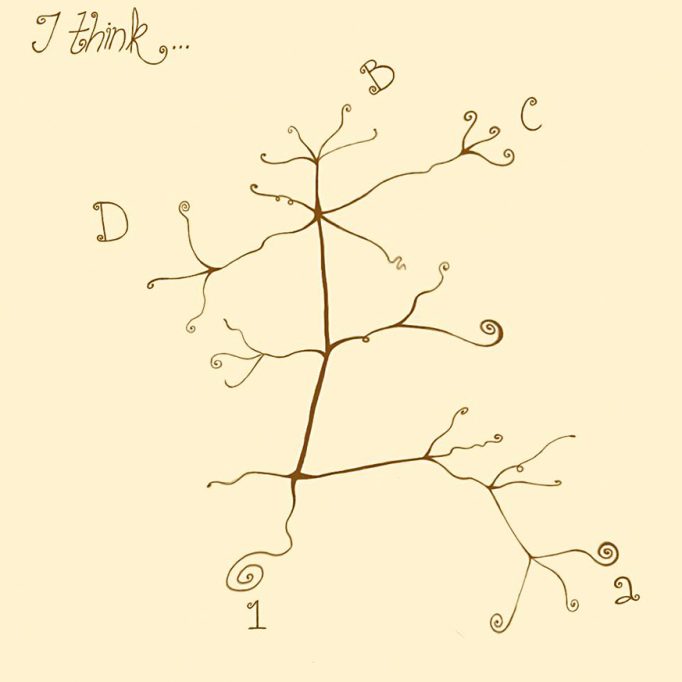
Figura 1. Darwin esquematizó la evolución de un grupo de especies a partir de una especie ancestral en su cuaderno de notas conocido como Notebook B. / Dibujo de Sofía Delaye Pascual
Como vemos, la teoría de la evolución de Darwin sugiere que las distintas especies que existen hoy en día han evolucionado (diversificado) unas de otras a partir de ancestros comunes. Esta idea, tan simple en principio, cambió profundamente la forma de entender a los seres vivos. Por ejemplo, cuando comparamos las extremidades de distintos mamíferos, encontramos que los huesos que las conforman son similares entre sí (figura 2). De acuerdo con Darwin, esta similitud se debe a que dichas extremidades evolucionaron a partir de un ancestro común, es decir, son homólogas. De hecho, Darwin escribió en El origen (1859, p. 415):
Como vemos, la teoría de la evolución de Darwin sugiere que las distintas especies que existen hoy en día han evolucionado (diversificado) unas de otras a partir de ancestros comunes. Esta idea, tan simple en principio, cambió profundamente la forma de entender a los seres vivos. Por ejemplo, cuando comparamos las extremidades de distintos mamíferos, encontramos que los huesos que las conforman son similares entre sí (figura 2). De acuerdo con Darwin, esta similitud se debe a que dichas extremidades evolucionaron a partir de un ancestro común, es decir, son homólogas. De hecho, Darwin escribió en El origen (1859, p. 415):
¿Qué puede haber más curioso que el que la mano del hombre, hecha para coger; la del topo, hecha para minar; la pata del caballo, la aleta de la marsopa y el ala de un murciélago, estén todas construidas según el mismo patrón […]?
El concepto de homología nos permite inferir parte de las características que poseía el ancestro común de un grupo determinado de organismos. Por ejemplo, en el caso de los mamíferos de la figura 2 podemos inferir que el ancestro común a todos ellos tenía estos huesos en las mismas posiciones relativas. La teoría de Darwin abre la posibilidad de que todos los seres vivos estemos relacionados en un gran árbol universal de la vida por haber evolucionado a partir de un único ancestro común. De hecho, Darwin sugirió en El origen (1859, p. 455): «[…] todos los seres orgánicos que en todo tiempo han vivido sobre la Tierra pueden haber descendido de alguna forma primordial.»
Sin embargo, Darwin, precavido como era en sus aseveraciones científicas, nunca se atrevió a representar la evolución de todos los seres vivos en un solo árbol. Fue Ernst Haeckel quien realizó uno de los primeros dibujos de un árbol universal. Como podemos apreciar en la figura 3, Haeckel sugiere que los seres vivos divergimos en plantas, animales y protistas a partir de un tronco común.
«La teoría de Darwin abre la posibilidad de que todos los seres vivos estemos relacionados en un gran árbol universal de la vida por haber evolucionado a partir de un único ancestro común»
Posteriormente, durante la primera mitad del siglo XX, Chatton (1938) y con mayor claridad Stanier y Van Niel (1941) propusieron que los seres vivos se deben de clasificar en dos grupos principales, a saber, procariontes y eucariontes (figura 4). Los eucariontes se definen por poseer membranas internas, tales como la membrana nuclear y el retículo endoplásmico, y por dividirse por mitosis, y los procariontes por carecer de membranas internas y dividirse por fisión binaria. En palabras de Stanier y Van Niel (1941, p. 464): «[…] el reino Monera, compuesto de microorganismos sin un verdadero núcleo, ni plastos, ni reproducción sexual.»
En 1969, el ecólogo Robert Whittaker propone que los seres vivos deben clasificarse en cuatro grandes reinos eucariontes (Plantae, Animalia, Protista y Fungi) y uno procarionte (Monera). Este esquema de clasificación fue adaptado por Lynn Margulis y combinado con la división procarionte/eucarionte para acomodar el origen de las mitocondrias y los cloroplastos a partir de las alfa proteobacterias y cianobacterias respectivamente (figura 5). Sin embargo, fue con el advenimiento de la biología molecular cuando la naturaleza del último ancestro común pudo comenzar a ser tema directo de investigación.
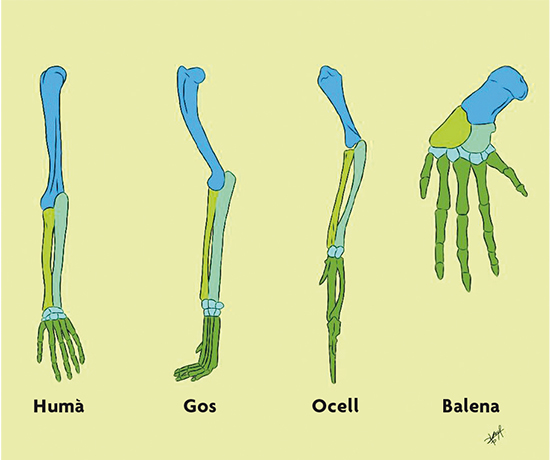
Figura 2. Extremidades homólogas entre distintos mamíferos. / Dibujo de Sofía Delaye Pascual
El ancestro universal, la historia del concepto
A mediados del siglo xx Frederick Sanger desarrolló las técnicas necesarias para conocer las secuencias de aminoácidos y nucleótidos que conforman a las proteínas y al ADN respectivamente. La información generada por esas técnicas permitió a Emile Zuckerkandl y Linus Pauling sugerir que era posible reconstruir la historia de los seres vivos comparando estas biomoléculas.
En 1977, utilizando técnicas moleculares, se publicó uno de los descubrimientos más sorprendentes sobre la diversidad de la vida en la Tierra. Carl Woese y George Fox descubrieron que, de acuerdo a las similitudes y diferencias en la subunidad pequeña de la molécula del ARN ribosomal (SSU rRNA por sus siglas en inglés), los seres vivos nos dividimos en tres grupos (Woese y Fox, 1977a). Estos tres grupos (o linajes) son: (a) las eubacterias, (b) el núcleo-citoplasma eucarionte, y (c) un grupo de procariontes al que denominaron como arqueobacterias. En la actualidad estos tres grupos se conocen como Bacterias, Eucariontes y Arqueas.
Basándose en esta división universal, Woese y Fox sugirieron que en la divergencia de los tres linajes celulares existía una entidad primitiva en donde la relación entre genotipo y fenotipo aún no había evolucionado a su forma actual (Woese y Fox, 1977b, p. 1). A esta entidad biológica ancestral la denominaron progenote: «Esta entidad primitiva se ha denominado progenote, término que muestra la posibilidad de que aún no se haya completado la evolución de la relación entre genotipo y fenotipo.» Algunos años más adelante, Fitch y Upper (1987, p. 761), al estudiar la evolución del código genético, acuñan el término cenancestro y lo definen como: «El antecesor común más reciente de todos los organismos que hoy están vivos.»
«Fue con el advenimiento de la biología molecular cuando la naturaleza del último ancestro común pudo comenzar a ser tema directo de investigación»
Es importante notar las diferencias entre ambos conceptos. El progenote implica un estado primitivo, en tanto que el concepto de cenancestro no necesariamente. El cenancestro, también conocido como «Last Universal Common Ancestor» o LUCA, es sólo el último ancestro común de todos los seres vivos que existen actualmente y este pudo haber sido simple como un progenote o complejo como una célula actual.
Gracias al desarrollo de la tecnología de secuenciación del ADN y a la acumulación de una gran cantidad y diversidad de secuencias en las bases de datos, fue posible comenzar a identificar los genes conservados entre las Bacterias, Arqueas y Eucariontes (figura 6). Ello permitió aplicar una metodología simple para inferir los genes que en principio se encontraban presentes en el genoma del cenancestro. Basados en esta metodología, Lazcano, Fox y Oró (1992) proponen que el último ancestro común poseía una maquinaria genética similar a la de una célula procarionte actual.
Sin embargo, la filogenia propuesta por Woese y Fox carece de raíz. Es decir, no muestra si alguno de los linajes celulares es más antiguo que los otros dos. La primera aproximación para tratar de identificar la posición de la raíz del árbol universal fue realizada independientemente por dos grupos de investigación al utilizar genes duplicados universalmente conservados. Ambos análisis sugirieron que la raíz del árbol universal se encuentra en la rama de las bacterias (figura 7). Ello reforzó la idea de que el cenancestro era de naturaleza procarionte.
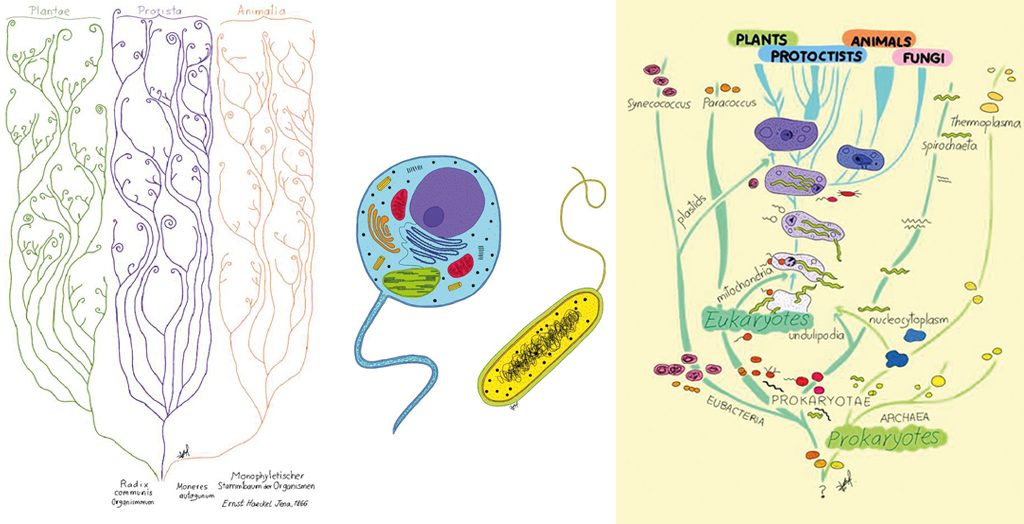
A la izquierda, Figura 3, árbol de la vida inspirado en Generelle Morphologie der Organismen (1866), de Ernst Haeckel. En el centro, Figura 4, observamos cómo a nivel celular podemos clasificar a todos los seres vivos en procariontes y en eucariontes. A la derecha, Figura 5, esquema de clasificación de los cinco reinos modificado por Lynn Margulis (1996). En la imagen podemos observar la división citológica de los seres vivos en procariontes y eucariontes. También podemos observar cómo los procariontes (Arqueas y Bacterias) dan origen a los eucariontes (Animales, Plantas, Hongos y Protistas) por simbiogénesis. / Dibujos de Sofía Delaye Pascual
Figura 6. Metodología utilizada por Lazcano, Fox y Oró para identificar genes presentes en el cenancestro. / Dibujo de Sofía Delaye Pascual
¿ARN o ADN?
En 1996 Mushegian y Koonin compararon los genomas de las bacterias Haemophilus influenzae y Mycoplasma genitalium para proponer el conjunto mínimo y suficiente de genes necesarios para sustentar la vida celular. También buscaron genes homólogos a H. influenzae y M. genitalium tanto en Arqueas como en Eucariontes para tratar de identificar genes universalmente conservados e inferir la naturaleza del último ancestro común. Dado que no encontraron genes homólogos para la enzima que se encarga de replicar el ADN en los tres linajes celulares, Mushegian y Koonin propusieron que el último ancestro común de los seres vivos poseía un genoma de ARN. Ello significaría que el último ancestro común sería más simple que cualquier célula conocida hoy en día. Sin embargo, inferencias posteriores han mostrado que el conjunto de genes conservados no es consistente con el nivel de complejidad de un progenote.
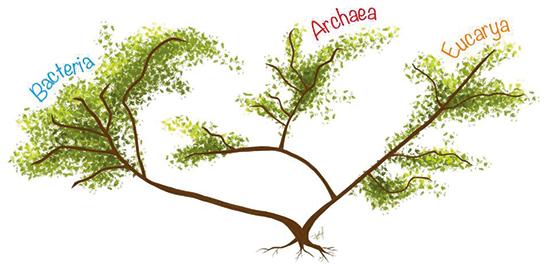
Figura 7. Representación del árbol universal basado en la filogenia de los tres dominios celulares con la raíz en el linaje de las bacterias. / Dibujo de Sofía Delaye Pascual
La transferencia horizontal de genes y el árbol de la vida
Una de las principales sorpresas que trajo consigo la secuenciación de genomas completos a finales de la década de los noventa fue la frecuente falta de concordancia de la filogenia propuesta por el ARNr, en donde los seres vivos se agrupan en Arqueas, Bacterias y Eucariontes, con las filogenias derivadas de otros genes conservados en estos tres linajes celulares. Si bien al inicio se pensó que muchas de las discrepancias se debían a problemas metodológicos, estudios posteriores demostraron que, al menos entre procariontes, una proporción importante de las incongruencias filogenéticas se debían al fenómeno de la transferencia horizontal de genes (figura 8).
El descubrimiento de la abundancia de eventos de transferencia horizontal a lo largo de la historia de la vida en la Tierra planteó un problema importante para la reconstrucción del conjunto de genes presentes en el último ancestro común. De hecho, se ha llegado a proponer que durante las etapas tempranas de la evolución de la vida en la Tierra la herencia horizontal era el modo más común de herencia (Woese, 1998).
Genes nuevos, genes perdidos y el cenancestro
Por un lado, un genoma puede ganar genes ya sea por transferencia horizontal o por creación de novo. Por otro lado, un gen que ya no contribuye a la supervivencia de un organismo puede perderse a lo largo de la evolución. Como ya vimos, la frecuencia de estos eventos afecta a la reconstrucción del contenido génico del último ancestro común. En un intento por tratar de inferir el contenido de genes del cenancestro tomando en cuenta los eventos de ganancia y pérdida de genes a lo largo de la historia de la vida en la Tierra, Mirkin y colaboradores (2003) desarrollaron un algoritmo basado en el argumento de la parsimonia. De acuerdo a su algoritmo, el último ancestro común tendría alrededor de 572 genes. Este conjunto de genes es casi suficiente para codificar un metabolismo coherente. Sin embargo, existen dos ausencias importantes. La primera de ellas, como ya mencionamos anteriormente, es la falta de una polimerasa de ADN. La segunda gran ausencia son dos enzimas clave encargadas de la síntesis de lípidos de la membrana celular.
La falta de estos componentes es sorprendente si tomamos en cuenta que representan dos de las tres propiedades más importantes de los seres vivos, a saber: la delimitación de un ser vivo de su ambiente a través de una membrana plasmática y la herencia del material genético basada en la replicación de un ácido nucleico (Szathmáry, 2005). La otra característica importante de los seres vivos, de la cual habla Eörs Szathmáry y que sí está presente en la reconstrucción de Mirkin es el metabolismo. La disputa sobre la naturaleza del genoma del último ancestro común y la naturaleza de sus membranas aún no está cerrada en la comunidad científica (Peretó, López-García y Moreira, 2004; Poole et al., 2014).
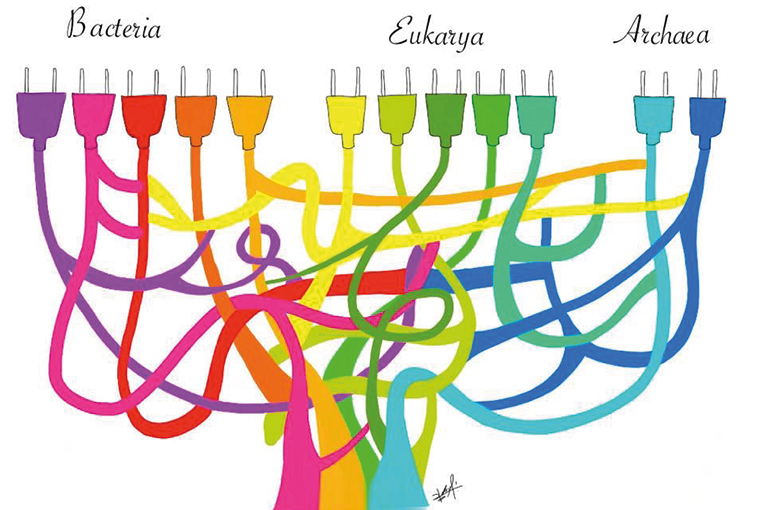
Figura 8. Representación del proceso de transferencia horizontal y anastomosis en el árbol universal. Las líneas de colores representan linajes celulares o genéticos. / Dibujo de Sofía Delaye Pascual
El último ancestro común y los virus
Otro aspecto no claro sobre la naturaleza del cenancestro es si los virus existían en su época y si este era susceptible a ser «infectado». Hasta el día de hoy no contamos con evidencia decisiva que manifieste la coexistencia de virus con células tan antiguas como el ancestro común. Sin embargo, diferentes hipótesis proponen que estas entidades no solo coincidieron, sino que los virus precedieron a las células, o incluso que tuvieron un papel central en etapas muy tempranas de la vida, cercanas a su origen (Agol, 2010).
Existen muchas preguntas sin resolver acerca del origen y la evolución de los virus. No obstante, muchos investigadores consideramos que estas entidades biológicas son de origen polifilético. Por ende, resulta muy difícil o imposible probar su presencia en etapas tempranas de la vida, debido a su naturaleza de material «fugado» de organismos celulares. Sin embargo, esta postura está lejos de despertar un consenso amplio y se enfrenta a una serie de objeciones. Por ejemplo, el hecho de que unas pocas, pero importantes, estructuras proteicas como el jelly-roll de la cápside esté presente en varios tipos virales, sugiere un origen común. Adicionalmente, se ha propuesto una lista pequeña de genes posiblemente homólogos a varios grupos de virus, los llamados hallmark genes (Koonin y Dolja, 2013), abonando la propuesta monofilética. De ser cierto, estos datos serían evidencia de un origen común para todos los virus, y sugeriría que dicho ancestro viral (o ancestros) fue contemporáneo o al menos cercano al ancestro celular.
Es importante recalcar que la información obtenida hasta el momento no permite discutir la presencia de los virus en épocas cercanas al cenacestro y no se puede extrapolar los datos obtenidos como evidencia de etapas más tempranas de la vida o mucho menos proponer que los virus tuvieron un papel central en el paso de organismos con genoma de ARN a ADN o incluso en el origen de la vida. Los métodos moleculares y la comparación de secuencias no pueden extraer información de etapas precelulares.
A pesar de esto, si el último ancestro común era ya un organismo tan complejo como una célula procarionte y si los virus son esencialmente material «fugado» de una maquinaria genética compleja, tampoco podemos descartar la presencia de virus en una época cercana al cenancestro. Saber si nuestro ancestro universal sufría infecciones virales o al menos coevolucionaba con ellos es algo que requiere de más estudios al igual que muchas de las preguntas que nos presenta el estudio de la evolución temprana de la vida.
Conclusiones
Hay un hilo que nos conecta desde nuestros días hasta los primeros seres vivos. Este hilo está formado por incontables generaciones de ancestros y descendientes modificados. Generaciones que en conjunto dan forma al árbol universal de la vida en la Tierra que alguna vez imaginó Darwin. Nuestras inferencias acerca de la biología del cenancestro están íntimamente ligadas a la estructura de este árbol universal y a los procesos evolutivos que le dan forma.
REFERÈNCIES
Agol, V. I. (2010). Which came first, the virus or the cell? Paleontological Journal, 44(7), 728–736. doi: 10.1134/S0031030110070038
Chatton, E. (1938). Titre et travaux scientifiques (1906–1937) de EdouardChatton. Sète: Sottano.
Darwin, Ch. (1859). On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. Londres: John Murray.
Fitch, W. M., & Upper, K. (1987). The phylogeny of tRNA sequences provides evidence of ambiguity reduction in the origin of the genetic code. Cold Spring Harbor Symposia on Quantitative Biology, 52, 759–767. doi: 10.1101/SQB.1987.052.01.085
Koonin, E. V., & Dolja, V. V. (2013). A virocentric perspective on the evolution of life. Current Opinion in Virology, 3, 546–557. doi: 10.1016/j.coviro.2013.06.008
Lazcano, A., Fox, G. E., & Oró, J. (1992). Life before DNA: The origin and evolution of early Archean cells. En R. P. Mortlock (Ed.). The evolution of metabolic function (pp. 237–295). Boca Raton: CRC Press.
Margulis, L. (1975). Symbiotic theory of the origin of eukaryotic organelles: Criteria for proof. Symposia of the Society for Experimental Biology, 29, 21–38.
Mirkin, B. G., Fenner, T. I., Galperin, M. Y., & Koonin, E. V. (2003). Algorithms for computing parsimonious evolutionary scenarios for genome evolution, the last universal common ancestor and dominance of horizontal gene transfer in the evolution of prokaryotes. BMC Evolutionary Biology, 3(2). doi: 10.1186/1471-2148-3-2
Peretó, J., López-García, P., & Moreira, D. (2004). Ancestral lipid biosynthesis and early membrane evolution. Trends in Biochemical Sciences, 29(9), 469–477. doi: 10.1016/j.tibs.2004.07.002
Poole, A. M., Horinouchi, N., Catchpole, R. J., Si, D., Hibi, M., Tanaka, K., & Ogawa, J. (2014). The case for an early biological origin of DNA. Journal of Molecular Evolution, 79 (5–6), 204–212. doi: 10.1007/s00239-014-9656-6
Stanier, R. Y., & Van Niel, C. B. (1941). The main outlines of bacterial classification. Journal of Bacteriology, 42(4), 437–463.
Szathmáry, E. (2005). Life: In search of the simplest cell. Nature, 433, 469-470. doi: 10.1038/433469a
Whittaker, R. H. (1969). New concepts of kingdoms of organisms. Science, 163, 150–160. doi: 10.1126/science.163.3863.150
Woese, C. R., & Fox, G. E. (1977a). Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proceedings of the National Academy of Sciences, 74, 5088–5090. doi: 10.1073/pnas.74.11.5088
Woese, C. R., & Fox, G. E. (1977b). The concept of cellular evolution. Journal of Molecular Evolution, 10, 1–6. doi: 0.1007/BF01796132
Woese, C. R. (1998). The universal ancestor. Proceedings of the National Academy of Sciences, 95, 6854–6859.




