


La cooperació és indispensable per a la vida. Les cèl·lules que componen el nostre cos i el de tots els animals, plantes i fongs troben el seu origen en diversos esdeveniments de fusió cooperativa (endosimbiosi) entre cèl·lules primitives, fa uns 2.000 milions d’anys. Tots i cadascun dels organismes multicel·lulars que trepitgen, naden o volen en aquest planeta, des dels diminuts àcars que s’alimenten de la pols que ens envolta a la colossal balena blava, són (som) resultat de la cooperació i integració entre cèl·lules que, originàriament, eren ens individuals de vida lliure. La més divertida de les formes de reproducció, la reproducció sexual, sorgeix també de la cooperació entre els sexes, i la pol·linització d’aproximadament el 90 % de les plantes amb flor, de la qual depenen els nostres ecosistemes, descansa sobre la cooperació entre plantes i distintes espècies d’insectes. En definitiva, les grans transicions evolutives, les societats animals i els nostres ecosistemes tal com els coneixem es fonamenten tots en l’existència de cooperació. No obstant això, en un món dominat per una competència feroç per la supervivència i la reproducció, l’existència de cooperació és, en si mateix, paradoxal, i la pregunta «per què cooperen els animals?» ressona avui amb més força que mai en biologia.
«Tots i cadascun dels organismes multicel·lulars que trepitgen, naden o volen en aquest planeta són resultat de la cooperació i integració entre cèl·lules que, originàriament, eren ens individuals de vida lliure»
En sentit evolutiu, la cooperació es defineix com una interacció entre dos organismes o entitats biològiques en què tant l’actor com el receptor resulten beneficiats, ja siga directament (per mitjà de la descendència pròpia) o indirectament (per mitjà de la descendència de familiars). És especialment enigmàtica la cooperació que es produeix entre dos organismes (o cèl·lules, o qualssevol entitats biològiques que interaccionen) que no sols no es troben emparentats entre si, sinó que ben sovint pertanyen a espècies distintes. La naturalesa està plena d’aquests casos, que denominem en termes generals mutualismes. Per exemple, els bacteris en l’intestí dels tèrmits que els permeten digerir cel·lulosa a canvi d’un ambient nutritiu on viure; els peixos netejadors que s’alimenten dels paràsits, mucus, crustacis o teixit mort de la pell dels seus clients per a benefici d’ambdós; o les formigues del gènere Atta, que metòdicament retallen i carreguen fulles fins al seu cau per alimentar els boscos de fongs que hi conreen, i que propagaran a altres colònies a canvi d’aliment. Però, com es mantenen aquestes aliances? En qualsevol dels exemples anteriors, què passaria si aparegueren individus «egoistes» que s’aprofitaren dels beneficis de la cooperació estalviant-se els costos d’haver de fer-la recíproca? Potser no li interessaria més a un peix netejador alimentar-se també del teixit sa del seu client? O a un bacteri de l’intestí d’un tèrmit deixar de contribuir a la digestió de cel·lulosa i aprofitar-se dels recursos metabòlics del seu hoste? Les aliances cooperatives permeten a ambdós actors accedir a recursos que altrament no estarien disponibles, per això probablement es troba tan estesa la cooperació, però com aconsegueix l’evolució conciliar el conflicte subjacent i evitar que les estratègies egoistes envaesquen i ensorren el sistema?
De tots els tipus d’aliances que trobem en la naturalesa, n’hi ha un que ha sigut i continua sent especialment útil per a abordar aquest tipus de qüestions. Es tracta de la coneguda associació entre formigues i pugons.
Pugons i formigues, una relació d’amor-odi
Els pugons són insectes xucladors que s’alimenten del floema (saba elaborada) de les plantes. El floema constitueix, a priori, una dieta desequilibrada per ser tan rica en sucres com pobra en aminoàcids (les «rajoles» a partir de les quals es construeixen les proteïnes i, per tant, part indispensable de la dieta d’un animal). Això fa que, per cobrir les seues necessitats d’aminoàcids, els pugons hagen d’ingerir grans quantitats de floema, la qual cosa genera un excés de sucres que és excretat per l’anus en forma de gotetes de melassa. La naturalesa rares vegades malbarata recursos tan preats com aquests, així doncs, les delicioses gotetes de melassa són aprofitades ben sovint per algunes espècies de formigues i això constitueix la base de les associacions de tipus mutualista descrites entre pugons i formigues (trofobiosi). Com vaques de sis potes, les formigues «munyen» la melassa dels pugons que, a canvi, reben protecció enfront dels depredadors i un exhaustiu «servei de neteja» per part de les formigues que els atenen, la qual cosa augmenta dràsticament les probabilitats de supervivència i reproducció de les colònies de pugons.
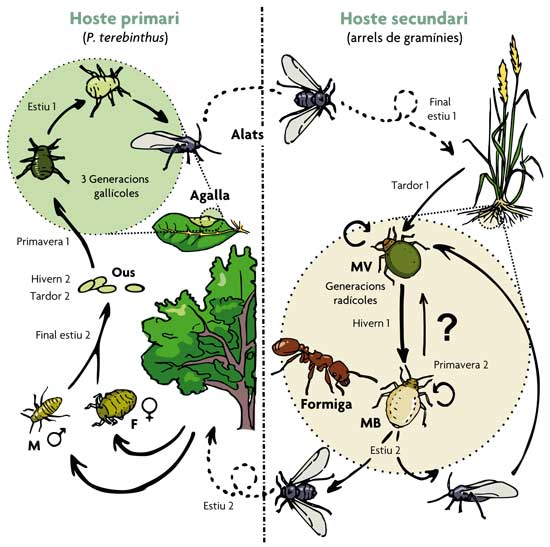
Figura 1. Esquema simplificat del cicle de vida bianual del pugó Paracletus cimiciformis. La fase sexual té lloc en l’hoste primari (arbustos de Pistacia terebinthus). Allí es produeixen fins a cinc morfos diferents. D’ells, tres són partenogenètics i es desenvolupen dins d’unes agalles molt característiques que indueixen en les fulles del seu amfitrió. Cap al final de l’estiu, l’última generació nascuda en l’interior de les agalles consisteix en pugons alats que volen i colonitzen les arrels de diverses espècies de gramínies (hoste secundari). Allí inicien una sèrie de generacions partenogenètiques àpteres (sense ales) formades per un o ambdós dels morfos alternatius: el morfo verd (MV) i el morfo blanc (MB). Aquests dos morfos interaccionen de manera molt distinta amb formigues del gènere Tetramorium. Al final de l’estiu, es produeixen dos morfos alats indistingibles que dispersen el clon a noves gramínies o tornen a Pistacia, donant lloc a una nova generació sexual. El signe d’interrogació indica que no es coneixen amb detall els factors que controlen la producció d’un o altre morfo. M, mascle; F, femella sexual. / Salazar, A., Fürstenau, B., Quero, C., Pérez-Hidalgo, N., Carazo, P., Font, E. & Martínez-Torres, D. (2015). Agressive mimicry coexists with mutualism in an aphid. PNAS, 112, 1101-1106. doi: 10.1073/pnas.1414061112
Encara que la descripció anterior constitueix el paradigma comunament utilitzat per presentar les interaccions entre pugons i formigues, la realitat, com quasi sempre, és més complexa. De fet, hi ha tot un ventall d’interaccions entre pugons i formigues que van des del mutualisme més estricte al més pur antagonisme. Així, algunes formigues combinen l’atenció dels pugons amb la seua depredació (és a dir, «cullen» tant la melassa produïda pels pugons que atenen com, eventualment i en alguns casos, els mateixos pugons), mentre que d’altres només són depredadores. Per la seua banda, de les aproximadament 4.000 espècies conegudes de pugons, només al voltant d’una quarta part participen en aquest tipus d’interaccions, mentre que la majoria no estableixen mai associacions amb formigues. A més, encara que hi ha nombrosos exemples de pugons mirmecòfils obligats (que només es poden desenvolupar sota la protecció de les formigues), la majoria d’associacions descrites entre pugons i formigues són de caràcter facultatiu (oportunista). En aquest sentit, diferents estudis sobre la distribució filogenètica de la mirmecofília en pugons suggereixen que les associacions entre pugons i formigues són evolutivament inestables, i que es formen i es trenquen en funció d’un complex balanç entre costos i beneficis per als participants que encara no comprenem bé. En definitiva, les interaccions pugó-formiga constitueixen una relació d’amor odi a escala evolutiva, i per tant un sistema ideal per a estudiar els mecanismes i els factors que regulen la cooperació.
«Les interaccions pugó-formiga constitueixen una relació d’amor-odi a escala evolutiva, i per tant un sistema ideal per a estudiar els mecanismes i els factors que regulen la cooperació»
Paracletus cimiciformis, dos pugons en un
Com la resta d’espècies del seu grup, Paracletus cimiciformis (el vertader protagonista d’aquest article) presenta un dels cicles biològics més complexos descrits en pugons (figura 1). Mentre que una part del seu cicle es desenvolupa sobre fulles de terebint (Pistacia terebinthus), una altra part té lloc sobre les arrels de diferents gramínies (fase radicícola) i representa una oportunitat per a establir contacte amb formigues i arribar a algun tipus de «acord» de cooperació. Durant més de 180 anys, des que es va descriure el 1837, només es va conèixer un únic tipus de pugó (el que anomenem un «morfo») sense ales per a la fase radicícola d’aquesta espècie: un morfo de coloració blanquinosa i amb forma de xinxa (d’ací el seu nom cimici, que en italià significa “xinxes”), del qual s’emfatitzava l’obligada associació amb formigues, principalment del gènere Tetramorium. No obstant això, el 2010 un estudi dut a terme en les universitats de València i Lleó (liderat per David Martínez, coautor d’aquest article) va demostrar que la fase radicícola d’aquesta espècie inclou un segon morfo sense ales, bombat i de color verd, que fins llavors havia estat classificat pels experts com una espècie diferent de pugó pertanyent a un altre gènere (figura 2). Aquest estudi va demostrar, a més, que els pugons d’ambdós morfos (verd i blanc) poden donar lloc a pugons del mateix o de l’altre morfo per reproducció asexual i que, per tant, són genèticament idèntics. A més de contribuir a millorar el nostre coneixement del cicle biològic de P. cimiciformis i resoldre un error taxonòmic arrossegat al llarg de quasi dos segles, aquesta troballa ens va obligar a plantejar-nos una pregunta òbvia: per què dos morfos?

Figura 2. Imatges de dos pugons adults corresponents al a) morfo blanc i b) al morfo verd de la fase radicícola de P. cimiciformis. A més de la diferent coloració, ambdós morfos es distingeixen en el nombre de peces de les seues antenes (sis en el morfo blanc i cinc en el verd) i en la forma de les seues tíbies posteriors (corbades i rectes, respectivament). / Carlos del Pico
Les dues cares de P. cimiciformis: Dr. Jekyll i Mr. Hyde
En un recent estudi multidisciplinari coordinat des de la Universitat de València (liderat per David Martínez i el seu estudiant de doctorat Adrián Salazar), ens vam proposar resoldre aquest enigma investigant amb detall les interaccions entre les formigues del gènere Tetramorium i els pugons dels morfos verd i blanc de P. cimiciformis. Els resultats –recollits en l’article «Aggressive mimicry coexists with mutualism in an aphid», publicat al gener de 2015 per la revista PNAS (Proceedings of the National Academy of Sciences of the United States of America)– van ser sorprenents i representen el descobriment d’una relació fins avui desconeguda entre pugons i formigues, i en general en la naturalesa: un sorprenent (i real!) cas de Dr. Jekyll i Mr. Hyde.
D’una banda, els pugons del morfo verd (el nostre particular Dr. Jekyll) mantenen amb les formigues una relació típica de trofobiosi (figura 3a). En contraposició, el morfo blanc es comporta com un autèntic Mr. Hyde. Quan les formigues s’aproximen als pugons blancs, s’inicia una sèrie d’esdeveniments ben diferents d’una seqüència trofobiòntica. Aquest particular ball comença amb les formigues palpant els pugons blancs suaument amb les antenes, com reconeixent-los, a la qual cosa aquests reaccionen retraient les seues potes i romanent immòbils. Llavors, les formigues els agafen amb atenció entre les mandíbules i els transporten a la seua cambra de cria, el lloc més protegit i impenetrable del formiguer, on els depositen junt amb les seues pròpies larves. Una vegada allí, els pugons blancs són netejats i atesos contínuament per les obreres de la mateixa manera que fan amb les seues pròpies larves. Tal és el grau d’integració dels pugons blancs en la cambra de cria de les formigues que costa distingir-los a simple vista de les larves de formiga que els envolten (figura 3b). Sense cap dubte, els pugons del morfo blanc són confosos per les formigues amb les seues pròpies larves, i el tracte que els dispensen resulta pràcticament indistingible del que tenen amb les seues larves. D’alguna manera, els pugons blancs imiten les larves de les formigues i aconsegueixen que aquestes els adopten (literalment).
«Hi ha tot un ventall d’interaccions entre pugons i formigues que van des del mutualisme més estricte al més pur antagonisme»
Per què? Quin és el sentit d’un morfo l’únic objectiu aparent del qual és fer-se transportar fins a la cambra de cria de les formigues? Les recurrents observacions de pugons blancs amb els seus estilets clavats en les turgents larves de les formigues no van deixar lloc a dubtes: el que aquests aparentment innocents pugons blancs fan en la cambra de cria és succionar l’hemolinfa (la sang dels insectes) de les larves de les formigues (figura 3c i 3d). Per tal de vèncer el nostre inicial (i sa) escepticisme, vam buscar DNA específic de formiga en l’interior dels pugons blancs amb accés i sense a la cambra de cria i només en vam trobar en aquells pugons que havien accedit a la cambra, deixant clar que aquests pugons «vampir» s’estaven nodrint de les larves de les mateixes formigues que els estaven adoptant. Això no sols representa el primer cas conegut d’un pugó carnívor (fins ara considerats estrictament vegetarians), sinó el primer cas en què, per mitjà de dos morfos (en aquest cas a més genèticament idèntics), una mateixa espècie es comporta alhora com a mutualista i com a depredador. Els dos morfos radicícoles (el cooperatiu morfo verd, que ofereix la seua melassa a les formigues, i l’agressiu morfo blanc, que enganya i xucla la sang de les larves de les formigues) representen així les dues cares d’una mateixa moneda, o els Dr. Jekyll i Mr. Hyde d’un mateix pugó: P. cimiciformis.
La pregunta següent: la base de l’engany
Una vegada descrit el fenomen, la pregunta que calia respondre era: «Com? Com aconsegueixen enganyar els pugons blancs a les formigues?». Se sap des de fa temps que el principal procediment de comunicació en les formigues és de tipus químic, gràcies a receptors presents en les antenes que els permeten detectar molècules de feromones o altres substàncies presents en l’aire o en el substrat. Més concretament, se sap que els hidrocarburs presents en la cutícula (la pell dels insectes) de les formigues representen un paper essencial en el reconeixement de membres de la seua mateixa espècie i colònia, i fins i tot en la distinció entre castes d’un mateix formiguer. Ens vam preguntar, per tant, si els pugons blancs no podrien estar imitant els hidrocarburs cuticulars de les larves de formiga, utilitzant-los com un «disfressa de corder» amb què ocultar la seua vertadera naturalesa de llop.
«En un món dominat per una competència feroç per la supervivència i la reproducció, l’existència de cooperació és, en si mateixa, paradoxal»
En una sèrie d’experiments realitzats en completa foscor, vam permetre a les formigues l’accés a una mescla de pugons del morfo blanc i del morfo verd de P. cimiciformis. Les formigues només van adoptar pugons del morfo blanc, per la qual cosa vam descartar que la discriminació entre ambdós morfos fóra visual. En una segona tanda d’experiments vam realitzar extractes de cutícula de pugons blancs, de pugons verds i de les mateixes larves de les formigues i els vam utilitzar per impregnar trossets de plàstic que després vam oferir a les formigues. Aquestes només es van portar a la seua cambra de cria aquells que havien estat impregnats amb extractes de les seues pròpies larves o de pugons del morfo blanc, la qual cosa apuntava clarament que les formigues confonen l’«olor» del morfo blanc de P. cimiciformis amb la de les seues pròpies larves. Finalment, una anàlisi química detallada, mitjançant cromatografia de gasos, va revelar que la composició en hidrocarburs cuticulars dels pugons del morfo blanc era, globalment, més semblant a la de les larves de les formigues que a la dels seus germans (genèticament idèntics!) del morfo verd, la qual cosa demostrava la naturalesa química del mimetisme desenvolupat pel morfo blanc de P. cimiciformis.
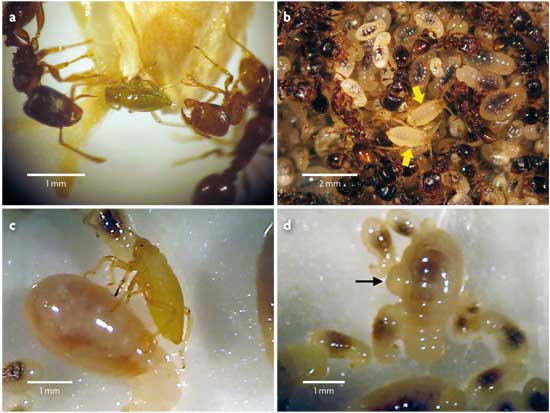
Figura 3. Imatges representatives de les interaccions entre els dos morfos de P. cimiciformis i formigues del gènere Tetramorium: a) interacció clàssica de tipus trofobiòntic entre el morfo verd (que ofereix la seua gota de melassa) i les formigues; b) resultat de l’adopció de pugons del morfo blanc per les formigues; la semblant coloració que presenta el morfo blanc i les larves de les formigues fa als pugons quasi indistingibles entre el muntó de larves de formiga; c) Mr. Hyde en acció: pugó blanc amb el seu estilet clavat en una larva de formiga, succionant la seua hemolinfa, i d) gota d’hemolinfa en una larva acabada de perfora. / Salazar, A., Fürstenau, B., Quero, C., Pérez-Hidalgo, N., Carazo, P., Font, E. & Martínez-Torres, D. (2015). Agressive mimicry coexists with mutualism in an aphid. PNAS, 112, 1101-1106. doi: 10.1073/pnas.1414061112
Més, i més, i més preguntes… El camí per recórrer
La descripció d’aquesta associació sense precedents entre pugons i formigues genera, sens dubte, una sèrie de noves preguntes a les quals caldrà anar donant resposta. Quins són els beneficis per al pugó P. cimiciformis de complicar, més si és possible, el seu cicle biològic introduint-hi un morfo agressiu? Com d’ampli és l’engany entre les distintes poblacions o espècies de formigues potencialment víctimes? Quin és el cost net de l’engany per a les formigues (convé tenir en compte que els pugons blancs produeixen eventualment pugons verds que ofereixen melassa a les formigues, i que per tant les beneficien)? Si el cost net és elevat per a les formigues, es troben pugons i formigues enmig d’una cursa evolutiva, les unes augmentant la seua capacitat de discriminació per a evitar l’engany i els altres «disfressant-se» cada vegada millor per a aconseguir els seus propòsits?
En tot cas, l’acabat de descobrir sistema P. cimiciformis-formigues es troba en la interfase mateixa entre la cooperació i l’engany i representa, per tant, una ocasió immillorable per a abordar l’estudi de nombrosos aspectes sobre l’origen i evolució de la cooperació i de l’explotació. A més, el fet sense precedents que dues estratègies evolutives ubicades en extrems oposats d’un continu teòric, que van des del mutualisme (el morfo verd) fins a l’explotació (el morfo blanc), coexistesquen en una mateixa espècie i en un mateix genotip és tan sorprenent com enigmàtic, tan nou com a excitant. És possible que altres aparentment innocents Dr. Jekyll tinguen allà fora en la naturalesa un malvat bessó Mr. Hyde?
Explicant l’altruisme: Hamilton i la selecció per parentiu
[El problema de l’altruisme és] especialment difícil, em semblava en un principi insuperable, i de fet fatal per a tota la meua teoria.
Charles Darwin, L’origen de les espècies, 1859
Els esquirols terrestres de Belding (Spermophilus beldingi) viuen en grups socials en les praderies de l’oest dels Estats Units. Quan un depredador s’acosta al seu grup social, de vegades un dels seus individus emet una vocalització o «crida d’alarma» que alerta els altres esquirols, que buscaran refugi ràpidament. Després de més de catorze anys d’observacions, Paul Sherman i els seus col·laboradors van poder calcular que les probabilitats que un esquirol que detecta un depredador terrestre siga capturat són d’aproximadament un 8 % quan aquest emet un senyal d’alarma, i tan sols d’un 4 % quan l’esquirol no emet cap senyal. Com ell mateix va reconèixer, aquest i molts més exemples semblants d’altruisme, en què un organisme sacrifica part de la seua eficàcia biològica en benefici d’un altre, van constituir una paradoxa central per a la teoria de la selecció natural de Darwin des del mateix instant en què es va publicar L’origen de les espècies el 1859. Com podia la selecció natural donar lloc a comportaments que disminuïen l’eficàcia biològica d’un individu?
«La descripció d’aquesta associació sense precedents entre pugons i formigues genera, sens dubte, una sèrie de noves preguntes a les quals caldrà anar donant resposta»
Hauríem d’esperar fins a 1964 per conèixer la resposta, quan William D. Hamilton va establir les bases de la «selecció per parentiu», una de les teories més reeixides de la biologia evolutiva. Utilitzant models matemàtics, Hamilton va posar de manifest que l’evolució pot afavorir aquests comportaments altruistes, i la cooperació en general, quan aquests tenen un efecte positiu sobre la descendència dels seus parents. Com que dos individus emparentats tendiran a portar els mateixos gens, un comportament altruista incrementarà de manera indirecta el nombre de còpies dels seus gens que es propaguen a la següent generació, i per tant pot resultar beneficiós en termes evolutius. D’ací la cèlebre frase del biòleg evolutiu J. B. S. Haldane, que, davant de la pregunta de si sacrificaria la seua vida per un germà, va contestar sense embuts que no dubtaria a sacrificar la seua vida sempre que fóra per la vida de dos germans, o vuit cosins. Com que dos germans comparteixen de mitjana un 50 % dels seus gens, un organisme «altruista» que sacrificara la seua pròpia descendència per la de dos germans (o vuit cosins, amb els quals comparteix el 12,5 % dels seus gens) tindria el mateix èxit reproductiu que un «egoista» que no se sacrificara. En sentit col·loquial, ambdós són igual d’egoistes, ja que ambdós estan maximitzant la seua pròpia eficàcia biològica, si bé per mitjà d’estratègies distintes.




