La immensa proliferació d’espècies que ha generat l’evolució ha estat i és causa d’estupor entre els científics. L’estupor no és un estat desitjable per a la ciència i no cal dir que molts han mirat de sortir-ne i donar respostes a la pregunta de per què n’hi ha tantes. Algun dels articles més famosos de la història de l’ecologia formula explícitament aquesta pregunta en el títol, com el cèlebre «Homenatge a Santa Rosalia» de Hutchinson. Però si és difícil de contestar en general, encara ho és més si considerem el cas específic de les plantes. Es coneixen de l’ordre de tres quarts de milió de plantes vasculars, i podem suposar raonablement que almenys una quantitat igual, si no molt superior, encara no ha estat descrita. Hi ha comunitats en què, en un sol metre quadrat de terreny, s’hi poden comptar fins a 60 espècies diferents de petites plantes, en bona part de vida efímera. Hi ha boscos tropicals que, en una hectàrea, contenen més de 300 espècies d’arbres, és a dir, que un de cada dos a cinc individus és d’una espècie diferent. El que fa particularment xocant aquesta diversitat vegetal és que totes les plantes empren quasi exactament els mateixos recursos. Els insectes poden menjar altres insectes, herba o fruits, fongs, xuclar saba o sang, i moltes més coses de moltes espècies de plantes i animals, però totes les plantes (llevat de les paràsites i carnívores) usen llum, diòxid de carboni, sals minerals i aigua, és a dir, uns recursos idèntics, pels quals han de competir. Com pot ser, doncs, que n’hi hagi tantes menes diferents? D’acord que l’ambient presenta certa heterogeneïtat, pendents amb sòls prims, fons de vall, afloraments de roques, solanes i obagues, etc., que poden oferir varietat d’ambients, però quan parlem de superfícies petites aquesta mena d’heterogeneïtat no pot ser massa important.

L’heura (Hedera helix). Arrapant-se als troncs, de vegades hi ha plantes enfiladisses que, amb menys inversió en fusta, poden també assolir el sostre del bosc i exposar les fulles a la llum. Foto: A. Aguilella
Endinsem-nos en un bosc dens. Si mirem en l’aire, veurem un dosser de fulles que quasi no deixa veure el cel i, de fet, només entre un u i un tres per cent de la llum que arriba a la cara exterior d’aquest dosser el travessa i assoleix el nivell dels nostres peus. Les fulles absorbeixen o reflecteixen la llum, n’aprofiten un u per cent o menys per fer la fotosíntesi, intercanvien oxigen i diòxid de carboni amb l’aire i s’aguanten allà dalt gràcies a les columnes de fusta dels troncs i les branques, per dins de les quals hi corren dos sistemes de canonades: una que puja aigua i sals, i l’altra que redistribueix els productes de la fotosíntesi. Arrapant-se als troncs, de vegades hi ha plantes enfiladisses que, amb menys inversió en fusta, poden també assolir el sostre del bosc i exposar les fulles a la llum. En boscos tropicals és freqüent que, a les enforcadures de branques o damunt d’aquestes, hi creixin plantes epífites, és a dir, que arrelen a la mica de terra acumulada en aquests indrets, sobre altres plantes, sovint a certa alçada. Als boscos atlàntics molt humits, pot haver-hi quantitats estimables de falgueres i molses també epífites. El nostre vesc (Viscum album) creix clavant-se per la base als teixits dels troncs dels arbres i viu d’una manera semiparasitària, fa la fotosíntesi, però xucla recursos de la saba. Hi ha, a més, arbres que no arriben al sostre, arbusts, mates, herbes, etc. Com més dens és el dosser, però, menys llum hi penetra i menys vida vegetal es desenvolupa per sota dels arbres dominants.
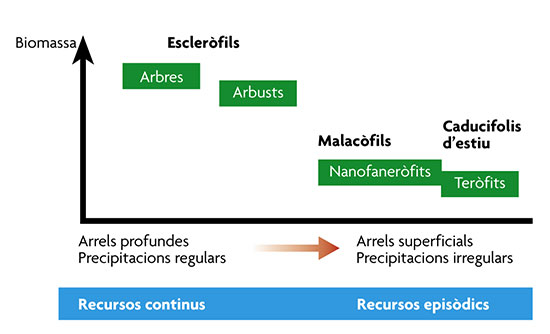
Tipus biològics de plantes mediterrànies en relació amb la disponibilitat de nutrients.
Per què no funciona sempre l’exclusió competitiva?
Els boscos mediterranis, de pi blanc, d’alzina, de surera, etc., sovint tenen una sola espècie d’arbre o, en tot cas, ben poques. En una hectàrea totalment coberta hi poden cabre des d’uns 600 individus (peus) d’arbres molt grans fins a uns 20.000 de petits, i ser tots de la mateixa espècie o, com a molt, de tres o quatre amb una de molt dominant. Com dèiem, però, en algunes selves tropicals la mateixa forma biològica, l’arbre, pot estar representada en igual superfície per centenars d’espècies. Això vol dir que costa una mica trobar un cert nombre d’individus de l’espècie que ens interessa, fet que ha tingut conseqüències pràctiques a l’hora de l’explotació. Per què conviuen tantes espècies d’arbres de semblant alçada que fan servir els mateixos recursos? No hi ha una llei ecològica anomenada d’exclusió competitiva, segons la qual, si dues espècies competeixen pel mateix recurs la que ho fa amb més eficiència exclou l’altra? Per què no s’han imposat les espècies que poden créixer més de pressa, estendre més aviat fulles i arrels i aprofitar amb més eficàcia aquests recursos, privant-ne les altres? Janzen va formular una possible explicació per als boscos tropicals. Potser tenir massa veïns de la mateixa espècie facilita la tasca d’expansió a fitòfags i plagues de tota mena, i l’evolució hauria seleccionat maneres de dispersió que portin les llavors a una certa distància, prudentment lluny, i això faria possible la coexistència de veïns diferents. La dispersió dels individus de la mateixa espècie té un cost, fa més difícil la reproducció, així que els avantatges deguts a la reducció de l’accés de paràsits i depredadors han de ser grans per fer versemblant aquesta hipòtesi.
«El que fa particularment xocant la diversitat vegetal és que totes les plantes empren quasi exactament els mateixos recursos»
Un ecosistema extremadament divers, aquest cop en espècies arbustives o mates, és el fynbos sud-africà, una mena de brolla. El fynbos creix sobre substrats pobres, amb un mosaic de condicions (proporcions relatives) de nutrients que pot haver afavorit la diversificació per heterogeneïtat de l’ambient. Hi ha una bona correlació entre la diversitat i l’escassesa de fòsfor. El règim de foc pot també tenir un paper ja que impedeix que hi hagi un procés d’exclusió competitiva, eliminant periòdicament els dominants. Tanmateix, la invasió recent per Pinus halepensis mostra que aquesta espècie suporta el règim de focs i tendeix a reduir seriosament la diversitat.

Aritjol (Smilax aspera). Foto: A. Aguilella
Un altre tipus d’ecosistema extremadament ric en espècies és el de les pastures mediterrànies. Des dels temps del professor salamanquí Fernando González Bernáldez, que va dirigir departaments d’ecologia a Sevilla i a Madrid els anys setanta i vuitanta, els ecòlegs de la seva escola van donar a conèixer les sorprenents diversitats de les pastures de la Meseta. Emprant l’índex de Shannon –que és una mesura treta de la teoria de la informació que es calcula a partir de la distribució dels individus en espècies, així que dóna 0 quan tots els individus són de la mateixa espècie i valors més alts quan hi ha moltes espècies i com més igualat és el nombre d’individus que pertanyen a cada una–, van trobar en aquelles comunitats de plantes valors que podien passar de 6 (valors propers a 5 ja són excepcionals en col·lectius d’un grup taxonòmic del tipus que sigui en una comunitat real). Als Monegres, quan començava la meva tesi doctoral, vaig veure aterrit com Oriol de Bolòs, el meu director, s’ajeia i pacientment feia un inventari d’unes quaranta espècies en un metre quadrat del que, de lluny, semblava un prat ressec i ras. Ací no eren arbres, sinó minúscules herbetes de pocs centímetres, com als prats de la Meseta. L’explicació de Janzen no sembla aplicable en individus que viuen uns mesos. Permeteu-me que em quedi amb el cas dels Monegres, que conec millor.

Lligabosc (Lonicera implex). Foto: A. Aguilella
Les comunitats més riques són als fons de la vall. Estan dominades per mates de gramínies perennes (Stipa spp., Lygaeum spartum, etc.) que s’obren per dalt com un ram però deixen molt d’espai al terra, on a la primavera poden sortir aquestes herbes efímeres. Part d’elles creixen també damunt els vessants dominats per matolls, però no hi solen assolir les mateixes riqueses. El nombre i identitat d’espècies i l’abundància de cadascuna varia segons l’any hagi estat més o menys plujós i fred. Aquests ambients contenen certa heterogeneïtat, per exemple més a prop de les mates de gramínies grans hi ha més ombra, la humitat i temperatura del sòl se’n veuen afectades de manera irregular. Hi ha pedres i petites elevacions o concavitats del terreny. Però hom pot sospitar raonablement que tanta diversitat no depèn només de les combinacions d’aquestes circumstàncies. La forta irregularitat interanual probablement fa que les condicions de vegades afavoreixin unes espècies, de vegades altres, així que l’abundància de cadascuna varia segons els anys, però la que enguany és una competidora superior l’any vinent pot no ser-ho. Alguns anys, poques arriben a germinar, altres ho fan quasi totes. D’altra banda, els herbívors suposen una pertorbació important, que inhibeix la competència. Per motius ben diferents que els arbres dels boscos tropicals, l’exclusió competitiva funciona malament en els sòls pobres i de condicions molt variables any per altre dels Monegres.
Llum i aigua: diferents estratègies de vida
«La llum és, doncs, un factor molt important d’heterogeneïtat espacial i temporal que permet diferents estratègies de vida»
Hi ha una creença ingènua, força arrelada entre els estudiants d’ecologia, segons la qual la major diversitat es dóna en les comunitats més madures. Els llibres d’ecologia expliquen que no sol ser així. Quan es tala o crema un bosc, s’inicia un procés de regeneració durant el qual el nombre d’espècies creix, en general fins que arbusts o arbres dominants ajunten les capçades. Llavors les herbes disminueixen. Com que hi ha més espècies d’herbes que no pas d’arbres, la successió continua amb un descens gradual del nombre d’espècies. Després, hom pot esperar que el bosc, si no és novament pertorbat, arribi a uns estadis de maduresa i envelliment durant els quals es produirà la caiguda ocasional d’alguns arbres per llamps o altres causes, obrint-se clarianes que produiran augments aïllats de diversitat amb nous descensos en tancar-se altre cop el dosser. La llum és un recurs necessari, i la varietat de plantes que existeixen s’adapten a determinades situacions en relació a la llum. Unes volen llum tot el dia, altres viuen millor a l’ombra, treballant a baixes intensitats de llum, altres aprofiten situacions intermèdies o fins i tot flaixos de llum provocats pel moviment de les fulles del dosser pel vent. L’aprofitament diferencial de la llum pot tenir lloc al llarg de l’any, com les espècies vernals que treuen les fulles molt aviat a la primavera, avançant-se a la foliació dels arbres caducifolis, posem per cas els faigs. Les anèmones que viuen sota els faigs acumulen reserves en bulbs subterranis i és gràcies a aquestes reserves que poden fabricar fulles abans que els faigs, fotosintetitzar i acumular noves reserves per a l’any vinent. La llum és, doncs, un factor molt important d’heterogeneïtat espacial i temporal que permet diferents estratègies de vida.
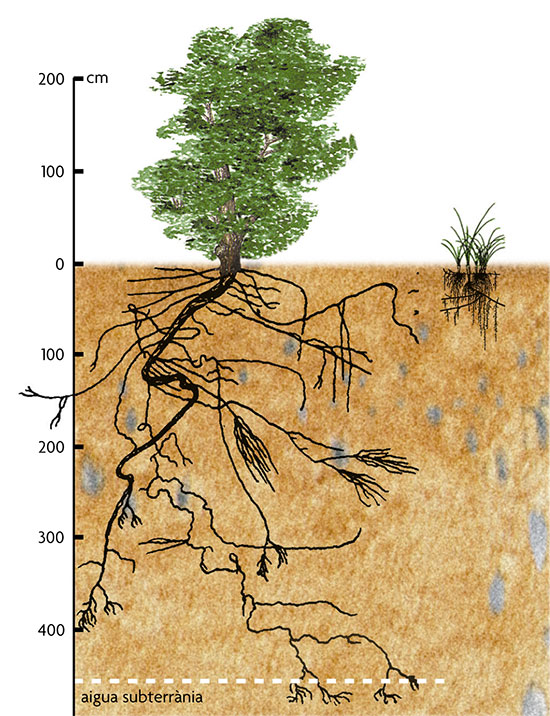
Dins un bosc, l’aigua no arriba de manera homogènia, hi ha punts de degoteig importants i molta aigua llisca pels troncs, humitejant el terra entorn seu. L’aigua en part s’infiltra en el sòl, i a certa fondària pot acumular-se de manera que la humitat hi és més constant. Les plantes amb arrels que arriben a aquesta fondària viuen en condicions relativament estables d’abastiment d’aigua i recursos, i poden mantenir les fulles hidratades i en activitat vegetativa durant força mesos. Les plantes d’arrels poc profundes només disposen d’aigua de manera irregular, com menys profundes més irregular és l’abastiment, i depenen de pluges puntuals. Solen tenir períodes vegetatius més curts i, en general, el seu ambient hídric és completament diferent del d’un arbre, encara que creixin a tocar, simplement perquè el subministrament dels arbres és molt més constant.
Un altre factor decisiu és l’aigua. Dins un bosc, l’aigua no arriba de manera homogènia, hi ha punts de degoteig importants i molta aigua llisca pels troncs, humitejant el terra entorn seu. Aquestes diferències espacials encara són més evidents en una devesa, ja que cada arbre forma una illa d’ombra, amb menys radiació i evaporació, aportacions de fullaraca amb nutrients, un cert efecte paraigua de la capçada, humitat al voltant del tronc, etc. Així que les mateixes plantes generen heterogeneïtat en la disponibilitat d’aigua, com en la de llum. L’aigua en part s’infiltra en el sòl, i a certa fondària pot acumular-se de manera que la humitat hi és més constant. Les plantes amb arrels que arriben a aquesta fondària viuen en condicions relativament estables d’abastiment d’aigua i recursos, i poden mantenir les fulles hidratades i en activitat vegetativa durant força mesos. Acostumen a tenir fulles amb contingut d’aigua poc variable (són hidroestables) i proteccions contra taxes excessives d’evapotranspiració com les cutícules gruixudes, pèls, etc. Les plantes d’arrels poc profundes només disposen d’aigua de manera irregular, com menys profundes més irregular és l’abastiment, i depenen de pluges puntuals. Les fulles suporten variacions importants del contingut d’aigua (són hidrolàbils), poden ser una mica suculentes, com el romaní, per ajudar a mantenir reserves, o tenen altres adaptacions que redueixen les pèrdues (fulles petites, peludes, estomes enfonsats en criptes, etc.), solen tenir períodes vegetatius més curts i, en general, el seu ambient hídric és completament diferent del d’un arbre, encara que creixin a tocar, simplement perquè el subministrament dels arbres és molt més constant. Les diferències de fondària de les arrels i de disponibilitat d’aigua s’associen a diferències en els nutrients. A les capes més fondes del sòl hi ha humitat més permanent, però pocs nutrients. Una alzina té, doncs, un abastiment força regular d’aigua i pobre en nutrients. Prop de la superfície, les arrels només tenen aigua quan plou, però els nutrients són abundants, així que en lloc d’un abastiment regular es tenen polsos bruscs d’aigua rica. La fondària de les arrels implica una gamma d’estratègies possibles en relació als recursos del sòl, com l’alçada i fenologia de les fulles ho fa pel recurs llum. Les pautes morfològiques del relleu també són causes d’heterogeneïtat que permeten diferents estratègies de vida.
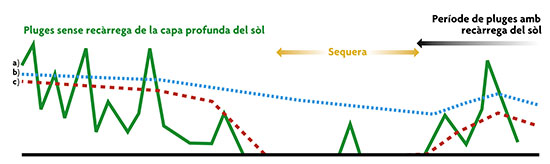
La variabilitat en la disponibilitat de recursos depèn de la profunditat de les arrels. En la figura es mostren les hipotètiques fluctuacions en la disponibilitat per a plantes a) d’arrels superficials; b) d’arrels profundes, en un medi en què les reserves profundes d’aigua no s’esgoten durant l’estiu; i c) en un altre en què les reserves profundes s’esgoten.
Tornem als Monegres. Allà trobem altres tipus de comunitats, a més de les de gramínies perennes i anuals efímeres. Cap d’elles s’acosta, ni de lluny, al nombre d’espècies de la que hem comentat. Hi ha una cosa, però, que crida l’atenció. A les comunitats de gramínies perennes i petites herbes anuals pràcticament només hi ha aquests dos tipus biològics de plantes, mentre que als matollars i boscos hi trobem més varietat, amb mates, arbusts, lianes, arbres i altres tipus d’herbes. Si apliquem l’índex de Shannon, al repartiment de la cobertura (o un altra mesura de l’abundància), en espècies, les comunitats de gramínies i anuals són molt més diverses. Si apliquem l’índex de Shannon a la distribució dels individus o la cobertura en tipus biològics (per exemple, emprant la tipologia clàssica de Raunkjaer), en canvi, la diversitat augmenta amb la complexitat estructural de la comunitat: els boscos tenen índexs de diversitat de tipus biològics més alts que els matollars i molt més alts que els prats. Això què vol dir? Que en els ecosistemes complexos hi ha més maneres diferents d’explotar els recursos, que aquests s’empren de manera més completa: els diferents nivells de llum, la disponibilitat més contínua o més episòdica d’aigua i nutrients, la fertilitat de capes profundes i superficials. I vol dir una altra cosa: que un ecosistema pot ser molt ric en espècies sense ser complex en l’aprofitament dels recursos. La riquesa en espècies no resulta de la complexitat de l’ecosistema, que es veu més reflectida per la varietat de tipus biològics, sinó que resulta d’inhibicions de diferents tipus a l’exclusió competitiva, com en els exemples que hem donat dels prats mediterranis i els boscos tropicals.
Aquesta idea s’allunya de les que acostumen a admetre ingènuament estudiants i, de vegades, textos o professors. És quan el medi biològic o físic imposa constriccions determinades que la riquesa d’espècies assoleix valors molt alts. Les plantes desmenteixen la llei de l’exclusió competitiva en la seva expressió més senzilla.
El pes de la història
«Un ecosistema pot ser molt ric en espècies sense ser complex en l’aprofitament dels recursos»
Si considerem la qüestió des d’una perspectiva evolutiva, és lícit preguntar-se per quin motiu, quan els vegetals van colonitzar la Terra (probablement, des dels inicis en simbiosi amb fongs per formar micorizes), no es formà una catifa més o menys contínua als llocs on hi havia humitat suficient. És probable que la primera fotosíntesi terrestre la fessin, en efecte, catifes de cianòfits i algues en llocs permanentment humits, però i a la resta? Una estructura en làmina, a l’estil de les hepàtiques, no funciona bé en l’intercanvi de gasos amb l’aire, ja que el fregament de l’aire crea sobre la làmina una capa d’aire quasi immòbil que es renova poc. Va resultar avantatjós, per tant, estendre’s en alçada, cosa que fou possible primer amb estructures que es mantenien dretes per turgència i després, molt millor, amb estructures rígides llenyoses que contenien vasos conductors. Aquestes estructures es van poder diversificar notablement, amb solucions que funcionaven millor sota les diferents condicions del medi i en relació també a la modificació del medi per les pròpies plantes. La fusta va permetre la diferenciació en alçada i prodigis com que els arbres més alts arribin als 120 m, és a dir, pugin aigua del sòl a aquesta alçada per abastir les fulles.

Foto: A. Aguilella
Les plantes llenyoses són més antigues que les herbàcies, en general. És a dir, dins de cada família de plantes, les llenyoses solen considerar-se primitives. Les primeres plantes terrestres aparegueren, segons sembla, fa al voltant de 400 milions d’anys. Aviat hi hagué falgueres arborescents i, als volts de fa 300 milions d’anys, coníferes. El seu èxit precedí els grans animals terrestres, inclosos els dinosauris, així que, a diferència del que passa al mar, on els animals consumeixen prop de la meitat de la producció de les plantes, la majoria de les quals no han desenvolupat estructures grans (fitoplàncton) a causa de la pressió del consum, en terra la fusta ha donat el domini dels ecosistemes a les plantes, i els animals no consumeixen enllà del 10% de la producció dels boscos i molts aprofiten, més que les plantes vives, la fullaraca i altres restes com a sapròfags.

El règim de foc pot també tenir un paper ja que impedeix que hi hagi un procés d’exclusió competitiva, eliminant periòdicament els dominants. Tanmateix, la invasió recent per Pinus halepensis mostra que aquesta espècie suporta el règim de focs i tendeix a reduir seriosament la diversitat. Foto: A. Aguilella
Els grans boscos acumulaven carboni i el retiraven de l’atmosfera. En part, anava a enterrar-se als sediments terrestres o, sobretot, marins. No estem segurs de les conseqüències d’això, però en tingué sobre el clima, que patí fluctuacions importants. Fem un salt en el temps, per aprofitar el que ens diuen els fòssils. El que no podien fer els animals, obrir les masses boscoses, ho féu en part el clima. Gràcies a importants canvis climàtics, es fa possible l’expansió de formes de vida herbàcia. El moment crucial es produeix fa entre setanta i cinquanta-cinc milions d’anys, al voltant de l’època en què s’extingiren els dinosauris, a finals del cretaci. Llavors es dóna la gran expansió de les gramínies i també comencen a imposar-se els mamífers, alliberats de la competència dels rèptils. Apareixen nous ecosistemes de tipus pastoral, amb espectaculars adaptacions entre plantes i animals, en què, ara sí, els animals adquireixen una preponderància que no tenien als boscos i poden assolir nivells de consum de la producció primària no massa diferents dels que hi ha al mar, mentre les herbes augmenten la relació entre producció i biomassa, així que aquesta es renova més de pressa que en els boscos. És notable que l’augment de les gramínies, que acumulen silici a les epidermis com una defensa contra els herbívors, té una correspondència força precisa amb l’expansió de les diatomees, que fan closques de silici, al mar. Segurament, l’activitat de les gramínies mobilitzà molt de silici que, en descompondre’s l’herba, fou arrossegat al mar i quedà disponible.
Conclusió
«Les primeres plantes terrestres aparegueren fa al voltant de 400 milions d’anys. Aviat hi hagué falgueres arborescents i, als volts de fa 300 milions d’anys, coníferes»
Em direu que he canviat de tema. No parlàvem de la diversitat de les plantes? Sí. El que pretenc demostrar és que la diversitat és possible 1) per l’heterogeneïtat ambiental; 2) per les diferents possibilitats d’explotar els mateixos recursos en el temps i l’espai; 3) per la creació d’heterogeneïtat pels organismes mateixos que fan ombra o ocupen espai; i 4) que tot això s’ha desenvolupat en un procés evolutiu històric en què han jugat la contingència en la seqüència d’esdeveniments, els canvis climàtics generals, els processos coevolutius entre organismes de tota mena i l’efecte de tot això en els cicles biogeoquímics, alliberant o retenint, segons els moments, quantitats enormes d’elements importants per a la vida. La diversitat general és fruit de tota aquesta història. La local és deguda a l’heterogeneïtat de condicions i a l’acció de mecanismes biològics o físics que impedeixen el domini d’una sola o poques «solucions» vegetals a l’ús dels recursos.