

Com s’adapten els organismes al canvi climàtic?
Les inversions cromosòmiques de Drosophila subobscura: el cas de les poblacions de Sèrbia

És conegut el fet que les inversions cromosòmiques de les poblacions naturals de l’espècie Drosophila subobscura són adaptatives respecte a les variacions de l’ambient. S’ha observat que les inversions canvien en freqüència al llarg del temps segons la predicció de l’escalfament global del nostre planeta. Aquests resultats es posen de manifest en els nostres estudis realitzats a poblacions sèrbies de D. subobscura. Així, el polimorfisme cromosòmic per inversions canvia segons les estacions de l’any i també a llarg termini (períodes de 10-15 anys) d’acord amb el que s’esperaria segons l’escalfament global. Per tant, les inversions cromosòmiques d’aquesta espècie són uns bons indicadors del canvi climàtic i també són útils per estudiar com els organismes s’hi poden adaptar.
Paraules clau: Drosophila subobscura, inversió cromosòmica, adaptació, selecció natural, canvi climàtic.
La genètica i el canvi climàtic
Els éssers vius tenen la capacitat d’adaptar-se als canvis ambientals. Les adaptacions no són més que unes característiques del disseny dels organismes que els hi permeten sobreviure i reproduir-se. Aquesta capacitat adaptativa, perquè sigui útil, ha de poder passar a la descendència, per tant ha de ser hereditària. És per aquesta raó que la genètica és tan important a l’hora d’explicar com afronten les espècies el repte d’un canvi ambiental tan intens i general com és el climàtic. Actualment coneixem que els genomes són uns grans reservoris de variabilitat genètica que està a disposició dels organismes per poder dur a terme les adaptacions i no desaparèixer.
«Les adaptacions són unes característiques del disseny dels organismes que els permeten sobreviure i reproduir-se»
El canvi climàtic actual representa una gran oportunitat per a la recerca, perquè podem aprendre molt sobre els mecanismes genètics de l’adaptació de les espècies. Aquests estudis es poden fer recollint mostres de la mateixa espècie al llarg del temps o en diferents indrets geogràfics (més freds o més càlids, per exemple) i veure quines diferències es troben per a alguns gens o marcadors genètics. Amb la informació d’aquestes observacions fetes a la natura es pot mirar de realitzar experiments al laboratori simulant condicions climàtiques particulars (normalment variant la temperatura) i analitzar si els organismes reaccionen igual respecte als mateixos gens o marcadors genètics. Aquests resultats poden donar-nos informació sobre la base genètica dels mecanismes adaptatius, en aquest cas, respecte al canvi climàtic. Actualment, i mercès a les anomenades tècniques de l’«ADN antic» podem fins i tot analitzar fragments del genoma d’organismes fòssils, tant d’espècies extingides com d’actuals, i analitzar quins d’aquests trossos del genoma poden ser clau en les adaptacions climàtiques. Tampoc podem oblidar que de vegades l’adaptació dels organismes no passa per tenir una variant o una altra d’un gen, sinó pels mecanismes de regulació d’aquests gens. La informació respecte a l’expressió gènica és molt valuosa perquè ens podria ajudar a preveure l’adaptació o no de poblacions d’organismes als canvis ambientals produïts per l’escalfament global.

D’altra banda, la genètica ens pot ser útil en l’altre sentit, en un vessant més pràctic. Si coneixem quins individus (o poblacions) presenten unes variants genètiques que els hi permeten sobreviure millor en unes determinades condicions ambientals es poden reconstruir alguns ecosistemes molt degradats pel canvi climàtic. Aquesta aproximació s’ha aplicat en la reconstrucció d’alguns ecosistemes dels Estats Units (Wymore et al., 2014). Aquesta tècnica passa per l’estudi genètic i ecològic de molts organismes, des dels bacteris fins a les plantes i els animals.
En aquest treball pretenem mostrar la nostra recerca sobre els mecanismes genètics de les espècies que els permeten adaptar-se, concretament al canvi climàtic. La nostra espècie és un tipus de mosca de la fruita, l’anomenada Drosophila subobscura, i els marcadors genètics que estudiem són les inversions cromosòmiques, és a dir, fragments de cromosomes que estan situats en una orientació oposada a l’habitual.
L’espècie: ‘Drosophila subobscura’
Drosophila subobscura és un dípter molt utilitzat en estudis de genètica, però sobretot d’evolució (figura 1). Té una distribució biogeogràfica (Krimbas, 1993) que abasta gran part d’Europa, excepte la part central i nord d’Escandinàvia i Islàndia. Al nord d’Àfrica es troba fins al desert del Sàhara i a l’Orient Mitjà és present fins l’Iran. La zona que ocupa als territoris de l’antiga Unió Soviètica és poc coneguda. També se la pot trobar a les illes de la Mediterrània, així com a les Açores, Madeira i Canàries. El 1978 es va descriure la seva presència per primer cop a Xile (Mestres, Balanyà, Prevosti i Serra, 1993), i es va expandir ràpidament per tota la regió a l’oest dels Andes, des de La Serena fins a Punta Arenas, malgrat que es creu que en la zona sud de Xile la seva presència podria ser estacional. Aquesta colonització segurament provindria d’una població mediterrània, però encara no se’n coneix amb exactitud l’origen. Uns anys després, en concret el 1982, es va trobar per primer cop a la costa oest de Nord-amèrica, des del sud de Califòrnia fins a la regió sud de l’estat canadenc de British Columbia. Cal esmentar també que un petit nucli colonitzador es va detectar al voltant de la ciutat argentina de Mar del Plata. Es pensa que seria una colonització secundària a partir de la principal de Xile. Hi ha una constant en les tres regions ocupades actualment per l’espècie, totes elles presenten la mateixa alternança de climes: marítim occidental i mediterrani (fins la zona semiàrida predesèrtica).
«Una inversió és un fragment de cromosoma que passa a col·locar-se en l’ordre oposat al normal. No representa cap problema per als individus que les presenten»
Una de les principals característiques de D. subobscura és que només té sis cromosomes politènics, que s’anomenen A (X o sexual), J, U, E, O i dot (puntiforme). Tots, a excepció del puntiforme, presenten inversions cromosòmiques (figura 2). Com ja s’ha comentat, una inversió no és més que un fragment de cromosoma que passa a col·locar-se en l’ordre oposat al normal. A D. subobscura les inversions s’anomenen per una lletra (que indica el cromosoma) i un número (que identifica la inversió concreta) o el símbol «st» (estàndard o referència). Molts cops un mateix cromosoma té diverses inversions formant el que s’anomena un «ordenament». Les diferents inversions que formen els ordenaments s’identifiquen pels seus números separats pel signe «+». Finalment, per mostrar si les inversions estan totalment o parcialment solapades es fa servir el subratllat (per exemple O3+4). El fet de tenir inversions no representa cap problema per als individus que les presenten, perquè tenen tots els mateixos gens, l’únic és que estan canviats d’ordre. En canvi aquest fet té una importància evolutiva crucial ja que els gens localitzats dins de la inversió tendiran a heretar-se junts. Si la combinació gènica que conté una inversió és favorable en el sentit que li permet adaptar-se millor al medi ambient on viu (és a dir, sobreviure i reproduir-se), aquesta serà afavorida per la selecció natural i per tant augmentarà en freqüència al llarg de les generacions. Per aquesta raó, D. subobscura ha tingut tant èxit com a espècie model en estudis evolutius.
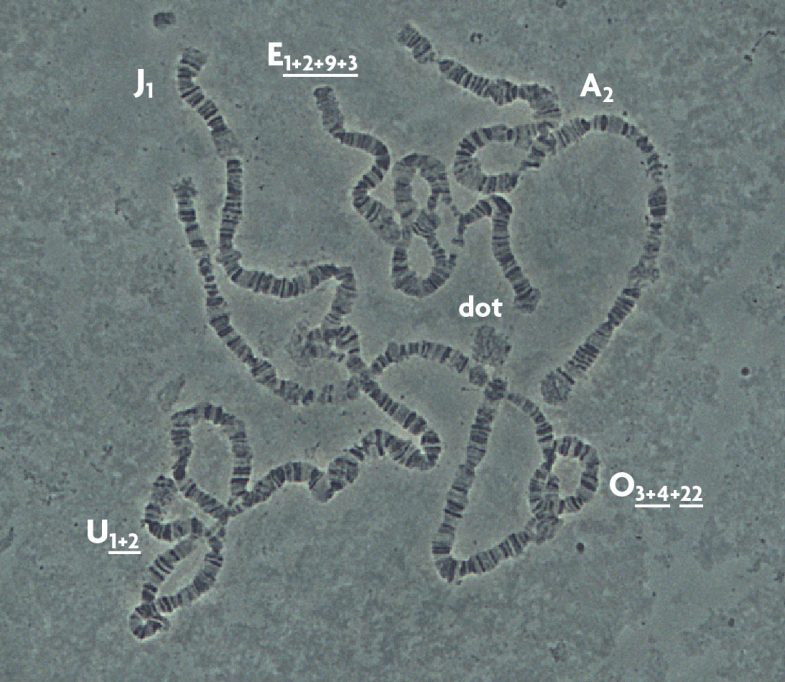
A partir dels anys cinquanta del segle passat diferents escoles europees d’investigadors començaren a analitzar el contingut de les poblacions naturals de D. subobscura respecte a les inversions. Aquestes presentaven variacions entre diferents poblacions i fins i tot entre diferents mostres d’una mateixa localitat. El que més va sorprendre aquells científics era que les freqüències d’algunes d’elles canviaven amb la latitud, és a dir, presentaven el que s’anomena un canvi clinal. D’aquesta manera, s’observava que unes inversions eren molt abundants al sud d’Europa i la seva freqüència anava disminuint en mostrejar més cap al nord (com per exemple O3+4). En canvi, d’altres que eren poc abundants al sud, anaven augmentant en freqüència en analitzar poblacions cap al nord (com el cas d’Ost). Hi ha dues explicacions del fenomen: una possibilitat seria que es tractés d’un procés històric. En un cert moment apareixeria una inversió (hi ha diferents mecanismes que les generen, però no els explicarem en aquest treball) i podria arribar a ser molt abundant en la població on apareix. Amb el moviment de les drosòfiles, la inversió s’aniria estenent passivament per Europa mostrant un gradient, i és clar, com més lluny del seu origen s’esperaria trobar-la en menor freqüència. La segona hipòtesi es basaria en el valor adaptatiu de les inversions. Una inversió nova, si aporta avantatge als individus que la presenten, augmentarà en freqüència mercès a la selecció natural. Si aquest avantatge és una adaptació a les condicions climàtiques (fonamentalment temperatura i d’altres factors lligats a aquest paràmetre), esperarem que la seva abundància depengui de la posició de les poblacions al llarg d’un eix nord-sud. Durant molts anys es va discutir quina de les dues hipòtesis explicava la distribució clinal de moltes de les inversions de l’espècie (Krimbas i Loukas, 1980). El fet de trobar a les dues regions colonitzades del continent americà (Xile i la costa oest dels Estats Units) clines latitudinals per a gairebé les mateixes inversions i amb el mateix sentit va donar un gran suport al valor adaptatiu de les inversions (Prevosti et al., 1988).
El canvi climàtic i les inversions cromosòmiques
Hi ha mesures meteorològiques que indiquen que la temperatura al nostre planeta va augmentant. Per exemple sembla que l’escalfament global és de 0,6 ºC a les tres dècades passades i de 0,8 ºC respecte al segle passat (Hansen et al., 2006). També la neu i el gel sembla que han disminuït vora d’un 10 % en el nostre planeta. En canvi, com a conseqüència, el patró de precipitacions ha esdevingut irregular. D’aquest fet se’n ressenten els éssers vius i els ecosistemes dels quals formen part. Per exemple, s’ha modificat el patró d’activitat estacional en molts organismes, així com també el temps de floració, de la caiguda de les fulles o de les migracions.
«Si la combinació gènica que conté una inversió és favorable perquè li permet adaptar-se millor al medi ambient, aquesta serà afavorida per la selecció natural»
Un dels primers investigadors a adonar-se de la importància del canvi de les freqüències de les inversions en el temps va ser el doctor Antoni Prevosti. Així, en un dels seus treballs (De Frutos i Prevosti, 1984) es van adonar tant de la presència de canvis estacionals com a llarg termini. Anys després, la darrera tesi que va dirigir, relacionà els canvis a llarg termini en les inversions cromosòmiques amb les variacions en la temperatura (Orengo i Prevosti, 1996). Una vegada més, Prevosti tornava a demostrar els efectes ràpids de la selecció natural a D. subobscura. Aquests treballs van tenir continuació de la mà del doctor Lluís Serra, que va analitzar el canvi a llarg termini de les inversions cromosòmiques en aquesta espècie comparant les dades antigues amb noves captures per una col·lecció amplia de poblacions europees (un total de tretze en un interval de quinze anys com a mínim) i també americanes, tant del nord com del sud (tretze poblacions analitzades en un interval entre divuit i vint anys). Si l’escalfament era global, a les poblacions europees, però també a les americanes, passats uns anys de l’origen de la doble colonització, les inversions haurien de canviar en freqüència segons les expectatives del canvi climàtic. En general, les poblacions analitzades tant a Europa com a Amèrica presentaven diferències a llarg termini tant en el tipus d’inversions com en les seves freqüències, de manera que les inversions considerades «càlides» anaven guanyant terreny amb el temps (Balanyà, Huey, Gilchrist i Serra, 2009).
Els estudis de les inversions cromosòmiques a Sèrbia
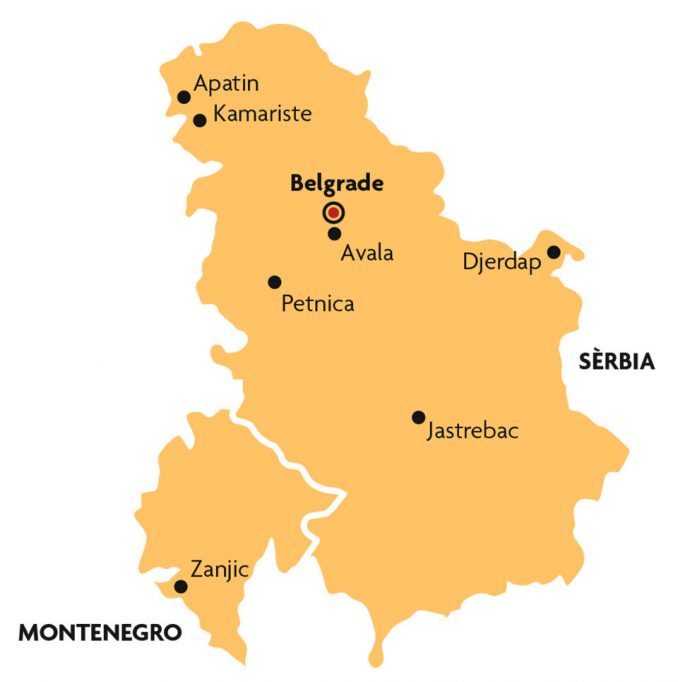
Per què era interessant esbrinar que passava a Sèrbia? Aquesta regió biogeogràfica era important per diferents raons. La majoria de clines latitudinals que històricament s’han estudiat a D. subobscura van en direcció sud-oest/nord-est, arrancant de la península Ibèrica. A més a més, encara que no es pot afirmar, hi ha tota una sèrie de proves genètiques que fan pensar que l’origen de la colonització d’Amèrica va tenir lloc a partir d’alguna població de la Mediterrània occidental. Per tant, era interessant veure si també les inversions canviaven i ho feien en el sentit predit per l’escalfament global en una altra regió europea diferent, en la qual no s’havia dut a terme cap estudi de clines latitudinals ni de variació al llarg dels anys. Les poblacions de D. subobscura dels Balcans tenen un contingut en inversions cromosòmiques que es pot qualificar de mediterrani, però és força diferent en alguns aspectes (tipus i freqüència d’algunes inversions) del que es troba en la conca occidental (Krimbas, 1993). Sèrbia, com que forma part dels Balcans, té com a avantatge ser considerada un refugi per moltes espècies durant la darrera glaciació. Aquest fet la fa evolutivament atractiva, ja que podria haver acumulat força variabilitat genètica. Les poblacions de D. subobscura d’aquest país han estat molt estudiades en matèria d’inversions, mercès a l’escola fundada pel doctor Dragoslav Marinkovic, i per tant es tenia molta informació d’anys enrere d’aquest marcador genètic (per a una bona revisió es pot consultar Krimbas, 1993 i Zivanovic, Andjelkovic i Marinkovic, 2002).

El primer estudi de la sèrie el vam realitzar a partir d’unes mostres de la població d’Avala, localitzada a 18 km de Belgrad (figura 3) i situada a uns 450 metres sobre el nivell del mar (Zivanovic i Mestres, 2010a). Es van fer captures durant els mesos de setembre de 2003, 2004 i 2005. Si es comparaven amb les mostres obtingudes a d’altres poblacions sèrbies, semblava que les freqüències de les inversions considerades «fredes» (en concret Jst, Ust, Est i Ost) havien disminuït, mentre que les d’algunes qualificades de «càlides» havien augmentat (J1, U1+2, U1+2+6, E1+2+9 i O3+4). També es va observar alguna petita variació entre anys consecutius. Això també és lògic, perquè les condicions ambientals dels mesos anteriors a la captura condicionen els resultats respecte a les inversions cromosòmiques. Cal recordar també que moltes de les inversions són adaptatives, no solament respecte a la temperatura, sinó a d’altres factors, que poden estar relacionats o no amb aquesta. En un altre treball, directament relacionat amb l’anterior, es van estudiar els canvis respecte a les estacions (Zivanovic i Mestres, 2010b). En concret es van comparar les inversions obtingudes al mateix lloc (Avala) però en una mostra de juny de 2004 respecte a una altra de setembre del mateix any. Es van trobar canvis per a algunes inversions (localitzades als cromosomes J, E i U) i en el mateix sentit que l’observat per altres investigadors.

Aquestes troballes ens van animar a continuar en aquesta línia de recerca, i per al següent estudi es va escollir la població d’Apatin, una zona humida del banc esquerre del riu Danubi (Zivanovic i Mestres, 2011). Es tracta d’un hàbitat força estable i poc afectat per l’activitat humana, idoni per a la recerca proposada. Es tenien dades dels cromosomes analitzats el 1994 i es van fer noves captures el 2008 i 2009. Una primera aproximació per veure si les inversions cromosòmiques varien respecte al que seria esperable segons el canvi climàtic és, com ja hem explicat, estudiar si les inversions considerades com a «càlides» i com a «fredes», respectivament, augmenten i disminueixen. Però per ser precisos cal estudiar si la temperatura canvia amb els anys. Per sort, vam poder obtenir dades de les temperatures màxima, mínima i mitjana, així com la pluviositat de la població, gràcies als registres del Servei Hidrometeorològic de la República de Sèrbia. Les temperatures mostraren una tendència a augmentar, mentre que el règim de pluges era irregular (fet també característic del canvi climàtic). Respecte a la composició en inversions, aquesta havia variat en el temps, detectant-se per primera vegada el 2009 l’ordenament U1+8+2, considerat «càlid». També van presentar un augment significatiu les inversions «càlides» U1+2, O3+4 i O3+4+22.
Posteriorment, es pensà d’estudiar els cromosomes d’una altra població però respecte a canvis mensuals (període curt de temps) i també en relació a mostres de feia quinze anys (llarg termini). La població sèrbia escollida va ser Petnica (captura realitzada l’any 2010), a uns 100 km al sud de Belgrad (Zivanovic, Arenas i Mestres, 2012) El lloc de captura era un bosc de Carpinus betula. En aquest treball també es va voler fer un estudi comparat mitjançant mètodes d’estadística multivariant de les poblacions sèrbies de les quals es tenien dades per les inversions cromosòmiques (un total de divuit mostres). Existien canvis mensuals en la composició cromosòmica de les mostres. Això pot ser degut al fet que la selecció natural actués en resposta als canvis ambientals que es van produint, però també al fet que els individus es poden desplaçar i poden buscar els llocs pels quals tinguin una preferència tèrmica. Quant als estudis a llarg termini, a la mostra de 2010 es van detectar per primer cop inversions «càlides» (U1+8+2, E1+2+9+12 i O3+4+8) no observades en la mostra de 1995. Una vegada més es va observar l’augment de les inversions «càlides» (A2, J1, U1+2 i O3+4) i la disminució de les «fredes» (Ast, Jst, Ust, Est i Ost). Un punt que cal destacar del treball és que a Petnica es va trobar un augment de totes les temperatures estudiades (màxima, mínima i mitjana), essent significatiu el canvi en la temperatura mínima.
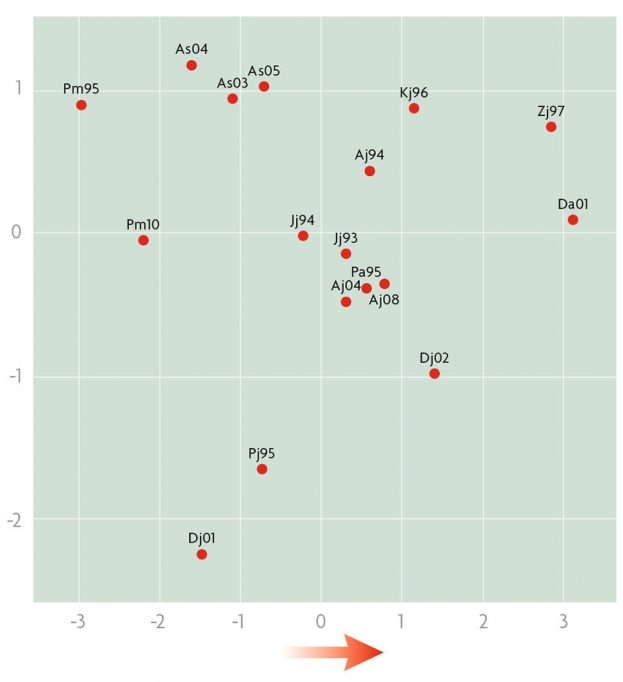
Quant a l’estudi de les inversions en el conjunt de les poblacions balcàniques, l’anàlisi clúster agrupava en general totes les «antigues» en un grup i totes les «recents» en un altre. Per tant, aquest marcador genètic es demostrava de nou que variava en el temps. A l’estudi de components principals, fent servir les dades de les inversions i de les característiques climàtiques de les poblacions, la primera i la segona component explicaven respectivament el 67,27 % i el 23,17 % de la variació total (figura 4). El que és interessant és que la primera explica les diferencies entre poblacions degudes a temperatura i pluges, és a dir, desplaçant-se respecte a ella en sentit positiu cada cop es troben poblacions més càlides i més seques. Per tant, la composició en inversions de les poblacions és una resposta de la selecció natural als canvis climàtics. És més, aquest canvi és en el sentit esperat per l’efecte de l’escalfament global del planeta. Aquest àmbit d’estudi és fascinant i la nostra intenció és continuar fent nous experiments i anàlisis, com per exemple veure l’efecte de les combinacions d’inversions en una mateixa parella d’homòlegs (cariotips) o les interaccions entre inversions de cromosomes de parelles diferents. Els nostres estudis preliminars semblen indicar que les inversions, per elles mateixes, tenen un valor adaptatiu més important que el dels cariotips (Zivanovic, Arenas i Mestres, 2014). En resum, les inversions cromosòmiques són un bon marcador del canvi climàtic i, a més a més, permeten estudiar de quina manera s’hi poden adaptar les espècies.
Referències
Balanyà, J., Huey, R. B., Gilchrist, G. W., & Serra, L. (2009). The chromosomal polymorphism of Drosophila subobscura: A microevolutionary weapon to monitor global change. Heredity, 103, 364–367. doi: 10.1038/hdy.2009.86
De Frutos, R., & Prevosti, A. (1984). Temporal changes of chromosomal polymorphism in natural populations of Drosophila subobscura. Genetica, 63, 181–187. doi: 10.1007/BF00128411
Hansen, J., Sato, M., Ruedy, R., Lo, K., Lea, D. W., & Medina-Elizade, M. (2006). Global temperature change. Proceedings National Academy of Sciences, 103, 14288–14293. doi: 10.1073/pnas.0606291103
Krimbas, C. B. (1993). Drosophila subobscura: Biology, genetics and inversion polymorphism. Hamburg: Verlag Dr Kovac.
Krimbas, C. B., & Loukas, M. (1980). The inversion polymorphism of Drosophila subobscura. Evolutionary Biology, 12, 163–234.
Mestres, F., Balanyà, J., Prevosti, A., & Serra, L. (1993). Genética evolutiva de la especie colonizadora Drosophila subobscura. Mundo Científico, 13, 408–416.
Orengo, D. J., & Prevosti, A. (1996). Temporal changes in chromosomal polymorphism of Drosophila subobscura related to climatic changes. Evolution, 50, 1346–1350. doi: 10.2307/2410676
Prevosti, A., Ribo, G., Serra, L., Aguade, M., Balaña, J., Monlcus, M., & Mestres, F. (1988). Colonization of America by Drosophila subobscura: Experimental in natural populations that supports the adaptive role of chromosomal-inversion polymorphism. Proceedings National Academy of Sciences, 85, 5597–5600. doi: 10.1073/pnas.85.15.5597
Wymore, A. S., Bothwell, H. M., Compson, Z. G., Lamit, L. J., Walker, F. M., Woolbright S. A., & Witham T. G. (2014). Community genetics applications for forest biodiversity and policy: Planning for the future. In T. Fenning (Ed.). Challenges and opportunities for the world’s forest in the 21st century (pp. 707–726) Londres: Springer.
Zivanovic, G., Andjelkovic, M. & Marinkovic, D. (2002). Chromosomal inversion polymorphism of Drosophila subobscura from South-eastern part of Europe. Journal of Zoological Systematics and Evolutionary Research, 40(4), 201–204. doi: 10.1046/j.1439-0469.2002.00189.x
Zivanovic, G., & Mestres, F. (2010a). Viabilities of Drosophila subobscura homo- and heterokaryotypes at optimal and stress temperatures. I. Analysis over several years. Hereditas, 147, 70–81. doi: 10.1111/j.1601-5223.2009.02163.x
Zivanovic, G. & Mestres, F. (2010b). Viabilities of Drosophila subobscura homo- and heterokaryotypes at optimal and stress temperatures. II. Seasonal component analysis. Hereditas, 147, 82–89. doi: 10.1111/j.1601-5223.2010.02164.x
Zivanovic, G., & Mestres, F. (2011). Changes in chromosomal polymorphism and global warming: The case of Drosophila subobscura from Apatin (Serbia). Genetics and Molecular Biology, 34, 489–495. doi: 10.1590/S1415-47572011000300020
Zivanovic, G., Arenas, C., & Mestres, F. (2012). Short- and long-term changes in chromosomal inversion polymorphism and global warming: Drosophila subobscura from the Balkans». Israel Journal of Ecology and Evolution, 58, 289–311. doi: 10.1560/IJEE.58.4.289
Zivanovic, G., Arenas, C., & Mestres, D. (2014). Inversion polymorphism in two serbian natural populations of Drosophila subobscura: Analysis of long-term changes. Russian Journal of Genetics, 50, 638–644. doi: 10.7868/S0016675814060150




