


La unicitat humana
La majoria de visions cosmogòniques sobre l’origen dels humans ens el mostren com a quelcom provinent de la divinitat, de fora del món natural. Això ha estat en part influït per la nostra evident unicitat. Perquè de totes les espècies d’humans del passat, potser més d’una desena, nosaltres som els únics que hem arribat fins a temps històrics. Si ens considerem excepcionals és, en el fons, per la constatació que som únics. Però si alguna altra espècie, com per exemple els neandertals, hagués sobreviscut, no hi ha dubte que el desenvolupament intel·lectual d’Occident hauria estat diferent.
Filòsofs com Plató, Aristòtil, Descartes, Rousseau, Hume o Marx han debatut sobre el vertader significat de la naturalesa humana, però les seves idees mai no han pogut ser contrastades amb el mètode científic, perquè els hi mancava una base empírica. En altres casos, s’han proposat com a definitoris dels humans alguns trets que després s’han trobat en altres primats, o fins i tot en mamífers no primats, com el reconeixement en mirall o l’existència de cultura o de tecnologia. Normalment es tracta de trets de tipus cognitiu o social (que són aspectes humans que nosaltres mateixos admirem) que han estat seleccionats de forma apriorística perquè encaixaven en una determinada visió ideològica de com havia de ser l’ésser humà. En aquesta llista, s’ha inclòs l’existència de llenguatge, d’art figuratiu, d’autoconsciència, d’una conducta simbòlica complexa, de la por a la mort i de la creença en un més enllà. Podem detectar la subjectivitat d’aquests plantejaments, per exemple, en el cas del llenguatge. L’evangeli de Sant Joan comença de manera impressionant amb aquesta afirmació: «en el principi era el Verb, i el Verb estava amb Déu, i el Verb era Déu».
«L’explicació més plausible és que aquestes regions cromosòmiques són el resultat d’una hibridació antiga amb neandertals que ha deixat petjades en els genomes no africans de la nostra espècie»
Hi ha gent, incloent-hi investigadors de psicologia evolutiva, que voldrien creure que aquest tret tan excepcional ha de ser exclusiu de la nostra espècie. Òbviament, ningú no estaria tan entusiasmat amb la idea definitòria proposada per l’escriptor rus Dostoievski, que deia, «només els humans poden ser tan artísticament cruels». Però la mateixa subjectivitat també afecta aquells que proposen trets que podem considerar com a negatius; s’ha descobert, per exemple, que altres primats també es dediquen a enganyar els seus congèneres per aconseguir menjar, sexe o atenció materna, i, per tant, tampoc la mentida és exclusivament humana. Per veure fins quin punt són problemàtiques totes aquestes definicions, només cal recordar que ningú no consideraria algú que fos mut, que no pogués crear art, que fos ateu o que fins i tot es trobés inconscient en un hospital, com si no fos humà. En aquest sentit, l’escriptor Italo Calvino no errava quan afirmava que «la raça humana és un conjunt d’individus que hauria de definir-se mitjançant la inclusió de tots ells». És a dir, es tractaria d’una aproximació a posteriori, contrastable des de la ciència: veure primer allò que tenim en comú tots els humans.
L’era de la genòmica ha obert un nou front per a estudiar la naturalesa humana. Disposem dels genomes de pòngids, els nostres parents vius més propers (alguns ja publicats, com el del ximpanzé, i d’altres, com el del bonobo i l’orangutan, en fase d’anàlisi), i tenim ja genomes humans personalitzats (cal recordar que el genoma humà del projecte original és en realitat un palimpsest de diferents individus), com els d’en Jim Watson i en Craig Venter. En el primer cas, però, és possible que es tracti de referències evolutives massa allunyades en el temps perquè puguin donar una informació molt precisa a nivell de gens amb una alta taxa de canvi evolutiu. És a dir, aquells gens que s’han modificat en el llinatge humà en les darreres desenes de milers d’anys, que és precisament quan sorgeix la nostra espècie. Novament, l’extinció de nombroses espècies d’hominí en els darrers dos milions d’anys, ha deixat un enorme buit d’informació evolutiva que l’estudi del genoma del ximpanzé no pot omplir.
Alguns investigadors creuen que si s’estudia al detall allò que compartim entre tots els genomes humans, podrem bastir una definició objectiva de la naturalesa humana. Però les primeres dades del projecte anomenat «dels mil genomes» estan començant a mostrar una altra dificultat: l’enorme variació que existeix en el genoma a nivell instraespecífic, fins i tot en trets, com les duplicacions de segments sencers de cromosomes, que fins ara no s’havien ni considerat. És a dir, podem trobar variants genètiques presents en tots els individus estudiats però no podem estar segurs que no hi manqui en el següent individu que analitzem. El problema serà especialment greu en les poblacions més variables de la humanitat actual, les de l’Àfrica sud-sahariana, com s’ha vist recentment en la publicació de diversos genomes khoisan i bantús. Una dificultat addicional seria la de categoritzar com a «exclusiu d’humà modern» variants gèniques que en realitat podrien haver estat presents en altres espècies extingides d’hominí. Això introduiria un biaix conceptual en la nostra comprensió de la naturalesa humana, perquè ens interessen precisament aquelles variants presents només en la nostra espècie.
«Els investigadors genòmics del projecte han descobert que en diverses seccions cromosòmiques, el genoma neandertal és més semblant als tres genomes no-africans que als dos africans»
Malgrat totes aquestes problemàtiques, és obvi que el que ens fa humans a tots per igual és el fet de compartir ancestres comuns, amb nombroses interconnexions que connecten tots els humans actuals en una gran xarxa evolutiva. Però això no significa que tothom comparteixi exactament tots els gens d’aquest procés. És a dir, el corrent de l’herència, la història evolutiva en comú és allò que ens acosta com a espècie, no un marcador concret en la humanitat actual. Per tant, si poguéssim accedir a la informació genètica d’una referència evolutiva més propera en el temps que la dels ximpanzés, llavors estaríem en la situació de poder delimitar exactament les novetats evolutives úniques de la nostra espècie.
El projecte genoma neandertal
Els neandertals (Homo neanderthalensis) haurien sorgit d’un seguit de poblacions més arcaiques del Plistocè mitjà europeu, sovint classificades com a Homo heidelbergensis. Van ser una espècie humana que va habitar Euràsia fa entre uns 400.000 i uns 30.000 anys, i que va iniciar el seu procés d’extinció amb l’arribada a Europa, fa uns 40.000 anys, dels nostres avantpassats procedents d’Àfrica. La naturalesa exacta de les seves interaccions durant aquest procés, el significat adaptatiu dels seus peculiars trets morfològics i l’abast real de les seves capacitats cognitives són encara motiu de controvèrsia entre els investigadors.

L’autor d’aquest article, Carles Lalueza-Fox forma part de l’equip multidisciplinar que estudia la cova asturiana i ha participat en el Projecte Genoma Neandertal.
© Equip d’Investigació d’El Sidrón
Les tècniques de biologia molecular han permès, des de 1997, la recuperació de tretze seqüències d’ADN mitocondrial (incloent-hi mitja dotzena de genomes mitocondrials o mitogenomes complets) de diferents neandertals, incloent-hi alguns del jaciment asturià d’El Sidrón, datats en fa 49.000 anys. L’ADN mitocondrial dels neandertals és diferent del dels humans actuals i molt semblant entre si, fins i tot entre individus separats per milers de quilòmetres. S’ha pogut estimar, mitjançant l’ús del rellotge molecular, que ambdós llinatges d’hominins es van separar fa mig milió d’anys. Tot això indica que els neandertals no varen contribuir a l’ADN mitocondrial dels humans moderns i que eren una espècie amb molt poca diversitat, i per tant, amb uns efectius demogràfics molt baixos.

L’investigador de la Universitat d’Oviedo Marco de la Rasilla i Svante Pääbo, l’investigador principal de l’Institut Max Planck, a la cova d’El Sidrón a Astúries. A més dels fragments de la cova de Vindija, l’esborrany es va complementar amb la seqüenciació d’altres tres neandertals procedents de Mezmaiskaia (Rússia), de Feldhofer (Alemanya) i d’El Sidrón.
© Equip d’Investigació d’El Sidrón
El Projecte Genoma Neandertal és un gran projecte científic liderat per Svante Pääbo, de l’Institut Max Planck d’Antropologia Evolutiva de Leipzig (Alemanya), portat a terme amb la col·laboració de la companyia privada 454 Life Sciences-Roche, al qual posteriorment s’ha afegit la companyia Solexa-Illumina. El projecte es va iniciar el juliol del 2006 amb l’objectiu d’aconseguir generar un esborrany genòmic neandertal en uns dos anys. La finalització d’aquest esborrany es va presentar públicament el 12 de febrer de 2009, en la simbòlica data de l’aniversari de Darwin. L’anàlisi de tots els milions de seqüències d’ADN va costar un any sencer, i la publicació no ha vist la llum fins el passat 7 de maig, a la revista Science.
La consecució d’aquest projecte ha estat possible gràcies a noves plataformes tecnològiques de seqüenciació massiva en paral·lel (també conegudes com a d’ultraseqüenciació) que no existien fa només quatre o cinc anys, i que han permès passar de l’artesanalitat de l’estudi de l’ADN antic a la generació de genomes extingits complets i del laboriós treball experimental a una anàlisi essencialment informàtica.
La major part de l’esborrany s’ha portat a terme a partir de tres petites mostres òssies del jaciment croat de Vindija, trobades l’any 1980 i etiquetades com a Vi33.16, Vi33.25 i Vi33.26. Les tres corresponen a individus femenins diferents, tot i que la primera i la darrera comparteixen el mateix ADN mitocondrial, i per tant, poden estar emparentades maternalment. L’esborrany generat té una cobertura genòmica de gairebé 2x, el que significa que, de mitjana, cada nucleòtid del genoma (i n’hi ha uns 3.100 milions!) és representat per dues seqüències. A la pràctica, però, això vol dir que un nucleòtid té una o dues seqüències per damunt i el veí potser no en té cap, perquè les seqüències es distribueixen a l’atzar. Per això tenim al voltant del 63% del genoma (cal afegir que les nombroses regions formades per seqüències repetides no es podran conèixer mai, donada la fragmentació de l’ADN original). L’esborrany de Vindija s’ha complementat amb la seqüenciació parcial de tres neandertals més, entre els quals hi ha El Sidrón 1253, provinent d’Astúries, i datat en fa 49.000 anys. En aquest darrer cas, s’han obtingut només 2,2 milions de nucleòtids, és a dir, un 0,1% del genoma.
Ni «Fora d’Àfrica» ni multiregionalisme, sinó tot el contrari
«Si hi ha hagut flux gènic, potser hem de dir que es tracta de la mateixa espècie, ja que el concepte biològic d’espècie és el d’una unitat reproductivament tancada»
Quan els analistes genòmics van començar a analitzar el genoma de Vindija i el van comparar amb els genomes complets de cinc humans actuals (dos africans –un ioruba i un san–, un europeu, un xinès i un nadiu de Papua-Nova Guinea) van descobrir un fet que els va deixar perplexos. I no només a ells; és evident que la troballa portarà a un canvi de paradigma en l’evolució humana. Jo també he hagut de modificar els meus plantejaments anteriors, fonamentats en evidències més limitades; però és així com funciona la ciència. El fet és que els resultats no s’ajusten a cap de les dues hipòtesis extremes i totalment incompatibles que s’han formulat sobre l’origen i evolució de la nostra espècie: la coneguda com a «Fora d’Àfrica» i la multiregionalista. Es tracta, doncs, d’una teoria nova de l’evolució humana.

Entrada de la cova de Vindija, a Croàcia. Els ossos utilitzats per elaborar l’esborrany genòmic del neandertal han estat trobats en aquest jaciment.
© Johannes Krause, Max Planck Institute for Evolutionary Anthropology
La hipòtesi «Fora d’Àfrica» (a vegades coneguda també com a Eva africana o Eva mitocondrial) manté que la nostra espècie es va originar a Àfrica fa entre 100.000 i 200.000 anys i que es va expandir posteriorment pels altres continents, alhora que reemplaçava les poblacions arcaiques locals (com els neandertals) sense creuar-s’hi. La majoria d’estudis genètics, i especialment els basats en el cromosoma Y i l’ADN mitocondrial, abonen aquesta hipòtesi, perquè les branques més basals d’aquests marcadors uniparentals sempre es troben a l’Àfrica sud-sahariana i perquè la diversitat trobada és molt baixa, i per tant, recent. Per contra, la hipòtesi multiregional, defensada per alguns morfòlegs que estudien fòssils, sosté que les poblacions humanes de cada continent deriven d’emigracions fora d’Àfrica molt antigues, properes al dos milions d’anys, per part d’hominins molt antics com els Homo erectus o formes similars. Aquestes poblacions haurien evolucionat localment, donant lloc als diferents grups humans de cada continent, però haurien mantingut una coherència morfològica i genètica (això és obvi, perquè tots els encreuaments humans donen descendents fèrtils) mitjançant el flux gènic, és a dir, el pas progressiu de gens d’una àrea geogràfica a una altra sense grans migracions, només per encreuaments minoritaris entre poblacions adjacents. A començament del segle xxi, la immensa majoria de científics –i tots els genetistes, podríem dir– defensaven la primera hipòtesi i de fet podria considerar-se que tot plegat era un debat ja tancat.
Doncs bé, els investigadors genòmics del projecte han descobert que, en diverses seccions cromosòmiques, el genoma neandertal s’assembla més als tres genomes no-africans (el de Nova Guinea, Europa i Xina) que als dos africans. Si la hipòtesi Fora d’Àfrica fos correcta, esperaríem just el contrari, perquè en estar els genomes africans situats més prop de la branca basal de tota la humanitat –i per tant, més a prop del tronc comú d’ambdós llinatges– haurien de ser els més semblants al genoma neandertal. De fet, això és el que s’observa en l’ADN mitocondrial. El mitocondrial humà més semblant al dels neandertals és precisament el de la hipotètica Eva mitocondrial africana, la mare de tots els mitocondrials humans.
«Els resultats del projecte científic no s’ajusten a cap de les dues hipòtesis que s’han formulat sobre l’origen i evolució de la nostra espècie. Es tracta d’una teoria nova de l’evolució humana»
En aquestes seccions de prop de 100.000 nucleòtids de llargada, presents en almenys 10 dels 23 cromosomes, la divergència evolutiva entre neandertals i no-africans és molt més recent que a la resta del genoma, perquè la seqüència d’ADN és pràcticament idèntica. Però quan es comparen les mateixes seccions amb les dels genomes africans, aquestes són molt diferents entre si, i per tant, donen temps de divergència (és a dir, de fondària evolutiva) més antics. No pot tractar-se de contaminació de fons (que en les mostres de Vindija ha estat estimada en un valor molt baix d’entre 0 i 0,5%), ja que aquesta es distribuiria uniformement per tot el genoma i no en forma de blocs discrets com es troba aquí. És a dir, si es contamina una mostra amb ADN extern, aquest es distribuirà per igual per tots els cromosomes. Per entendre tot això, imaginem-nos que som el resultat d’una hibridació; el nostre cromosoma 16, per exemple, ha rebut una còpia del pare (posem neandertal) i una de la mare (humana moderna), i cada còpia és diferent en parts de la seva seqüència. Quan nosaltres tenim descendència, no passem a aquesta una o altra còpia dels nostres cromosomes 16, sinó que en generem un de nou que és un palimpsest de les dues còpies que ja tenim (per un fenomen d’intercanvi genètic que es coneix com a recombinació). És a dir, el cromosoma 16 de la següent generació tindrà un bloc del 16 patern, després un bloc del 16 matern, etc. A mesura que ens allunyem en el temps d’aquesta hibridació puntual, els blocs s’aniran fragmentant per successives recombinacions en els descendents, però així i tot continuaran essent discernibles en una anàlisi genòmica global. Això és el que es troba en els genomes neandertals i dels humans no-africans estudiats, i representa entre l’1 i el 4% dels genomes d’aquests darrers.

Els investigadors van obtenir la majoria de l’ADN utilitzat per al seu estudi d’aquests fragments d’os pertanyents a tres femelles neandertals que van ser trobades a la cova de Vindija a Croàcia.
© Max Planck Institute for Evolutionary Anthropology
L’explicació més plausible és que aquestes regions cromosòmiques són el resultat d’una hibridació antiga amb neandertals que ha deixat petjades en els genomes no africans de la nostra espècie. Perquè això s’hagi pogut produir, el fenomen ha d’haver tingut lloc poc després de la sortida comuna d’Àfrica, fa uns 100.000 anys. La petjada genòmica no és prou recent perquè els darrers neandertals s’hagin creuat amb els humans moderns que entren a Europa fa 40.000 anys. Entre altres coses, si hagués estat així, el rastre evolutiu només es trobaria en l’individu europeu, i no en els tres no africans, que n’inclouen un d’Oceania.
On va tenir lloc, doncs, aquest increïble encontre? Des de fa molts anys se sap que neandertals i humans moderns primitius van coexistir a la zona del Proper Orient (i probablement per tot l’Orient Mitjà). A Israel, trobem jaciments molt propers que contenen la mateixa tecnologia lítica del paleolític mitjà però amb restes fòssils que són o bé neandertals o bé humanes modernes. Les datacions, però, no són coincidents i això havia fet pensar que, malgrat la proximitat geogràfica, potser mai no es van trobar en persona. El neandertal arcaic de Tabun té una datació de 120.000 anys, els humans moderns arcaitzants de Skhul i Qafzeh al voltant de 90.000-100.000, i els neandertals de Kebara i Amud al voltant de 60.000. A més a més, hi ha altres restes neandertals a Síria (Dederiyeh) i a Iraq (Shanidar). És probable que en tota aquesta regió hi hagués alguns encreuaments quan els nostres avantpassats van començar a sortir d’Àfrica. Quan una població colonitzadora com la dels humans moderns es troba amb una població resident, fins i tot un nombre molt petit d’encreuaments entre ambdues poblacions pot deixar una petjada genètica força gran en la primera, per la mateixa dinàmica expansiva de la població colonitzadora. Com a conseqüència, el flux gènic normalment només es detecta, com és el cas, de la població resident a la colonitzadora. El fet que els llinatges mitocondrials siguin clarament diferents pot informar-nos d’un biaix sexual en aquests encreuaments (homes neandertals amb dones humanes modernes), però de fet també seria compatible amb la pèrdua per efecte de l’atzar al llarg de les generacions dels mitocondrials d’origen neandertal si el flux gènic va ser, com sembla, limitat
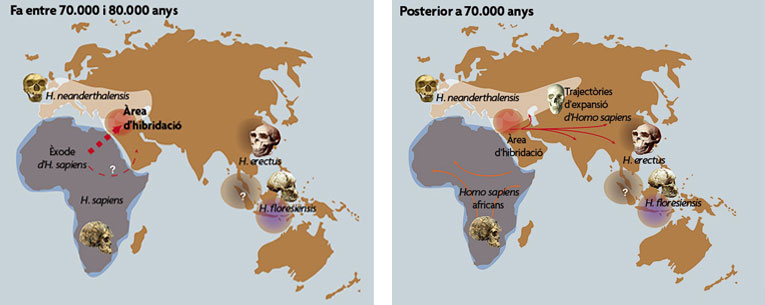
Aquests mapes mostren com es va produir la interacció entre humans moderns i neandertals ara fa entre 70.000 i 80.000 anys. Segons la investigació realitzada per investigadors de l’Institut Max Planck en col·laboració amb científics espanyols del CSIC, aquesta hibridació podria haver tingut lloc al Proper Orient (mapa de l’esquerra), i des d’allí s’hauria expandit per Euràsia i Àsia, com es veu en el mapa de la dreta.
© Comunicación CSIC / Antonio Rosas
Finalment, hi ha altres zones en el genoma neandertal que mostren una profunditat evolutiva més gran que la resta. Seguint el mateix raonament, correspondrien probablement a rastres més antics d’hibridacions amb humans encara més arcaics. Òbviament, no disposem del genoma d’un Homo erectus o d’un Homo antecessor per a poder mirar si comparteixen aquestes zones genòmiques, però tot sembla indicar que diferents expansions humanes han comportat que hi hagués repetidament flux gènic amb altres grups procedents d’expansions anteriors.
Un estudi publicat recentment apunta també en aquesta direcció. Es tracta de la recuperació del genoma mitocondrial d’un petit os del dit de la mà trobat a la cova siberiana de Denisova i datat entre fa uns 30.000 i uns 48.000 anys. La seqüència mitocondrial correspon a un llinatge que és molt diferent dels humans moderns i també dels neandertals, i el temps de divergència d’aquest ha estat estimat en un milió d’anys. Tot i que els periodistes van interpretar això com la troballa genètica d’una nova espècie, cal recordar que no hi ha una correspondència directa entre divergència genètica i diferència d’espècie. Algunes espècies estan estructurades geogràficament i això provoca notables diferències evolutives entre individus. Per exemple, se sap que entre els mamuts hi havia dos grups de llinatges mitocondrials que havien divergit fa entre 1,5 i 2 milions d’anys, i ningú no ha dit que fossin dues espècies diferents de mamut. Podria ser que Denisova fos, genòmicament, morfològicament, i també culturalment, un altre neandertal, però que el seu llinatge mitocondrial fos el rastre d’un flux gènic amb hominins anteriors. Òbviament, això es podrà saber estudiant el seu genoma nuclear, ja que la forma peculiar de transmissió de l’ADN mitocondrial ens emmascara les possibles explicacions evolutives.
Una dificultat inesperada
«Per a comprendre el que representen els canvis trobats, haurem d’estudiar-los un a un, i entendre’n la funció diferencial en humans moderns i en neandertals»
Ara bé, tot això comporta una nova dificultat. Per saber allò que és exclusivament humà necessitem disposar d’una referència evolutiva externa, però si hi ha evidències de transferència de gens entre els llinatges dels neandertals i dels humans moderns, potser no la tenim. És a dir, si hi ha hagut flux gènic, potser hem de dir que es tracta de la mateixa espècie, ja que el concepte biològic d’espècie és el d’una unitat reproductivament tancada. A nivell genètic, no hi ha res que permeti definir que dues poblacions (o dos individus) són d’espècies diferents; uns pocs canvis genètics poden comportar barreres reproductives. A nivell morfològic tampoc; coneixem casos d’organismes –les anomenades espècies críptiques–, que són gairebé indistingibles externament i en canvi pertanyen a espècies diferents. Però fet i fet, el concepte d’espècie no deixa de ser una idea convenient, un concepte, diguem-ne, operatiu, que necessitem els biòlegs per treballar, des dels taxònoms fins als conservacionistes. El mateix Darwin creia que els llinatges evolutius eren un continuum i que tallar-los en espècies era una arbitrarietat nostra. Tenia raó; per exemple, és absurd considerar que, en algun punt del llinatge neandertal, trobarem un individu que serà ja neandertal mentre que els seus progenitors seran encara Homo heidelbergensis.

Diversos cranis de neandertals i al fons un d’Homo sapiens. La publicació del primer esborrany del genoma neandertal ha revelat que Homo sapiens eurasiàtic comparteix d’un 1% a un 4% del seu ADN amb els neandertals.
© Comunicación CSIC
És clar que diverses evidències semblen indicar que es tractaria de dues espècies diferents: per exemple, la morfologia neandertal, tan característica, es troba fora del rang de variació de la humanitat actual, i diversos marcadors genètics, com el ja mencionat ADN mitocondrial, mostren una clara separació entre ambdós llinatges. El més plausible és pensar que va haver-hi un fenomen restringit d’hibridació quan les dues espècies van entrar en contacte fa potser 100.000 anys. Aquesta troballa inesperada no invalida, tot i que dificulta, el nostre plantejament inicial per caracteritzar la naturalesa humana.
La definició genètica d’ésser humà
La forma més directa de buscar singularitats en el genoma neandertal és buscar aquells gens que presentin canvis d’aminoàcid, que, amb tota probabilitat, afectaran la funció de la proteïna codificada. S’han localitzat 78 gens amb aquests tipus de canvis, tot i que a causa de la baixa cobertura genòmica n’hi podria haver fins a tres cops més. Alguns gens de la llista són fàcils d’interpretar. Per exemple, hi ha canvis en cinc receptors olfactius que nosaltres probablement tenim inactivats perquè avui dia no són importants per a la nostra supervivència. Nosaltres, més que cap altre primat, som animals essencialment visuals.
En cinc gens més hi ha dos canvis d’aminoàcid (la qual cosa implica un canvi de funció encara més important). Tenim el gen SPAG17, que sembla tenir un paper important en el moviment de l’esperma; el gen TTF1, que és un factor de transcripció que actua activant altres gens; el gen DCHS–1, que codifica per a una proteïna que intervé en la cicatrització de ferides; el gen RPTN que codifica per a una proteïna que s’expressa a l’epidermis i que està implicada en les glàndules sudorípares, la rel dels cabells i les papil·les de la llengua, i el gen SOLH, que codifica per a una proteïna que encara no sabem quina funció té.

L’investigador Svante Pääbo, amb la reconstrucció d’un crani neandertal. El científic de l’Institut Max Planck, un dels més reconeguts experts en ADN fòssil, ha liderat el Projecte Genoma Neandertal, del qual es presentaren les conclusions al maig de 2010.
© Frank Vinken
També s’han localitzat 82 regions que mostren senyals d’haver estat seleccionades de manera positiva per les forces evolutives (això es detecta perquè aquest procés deixa una zona de baixa diversitat genètica al voltant). En una d’aquestes regions hi ha el gen AUTS2, que codifica per a una proteïna que s’expressa en el cervell durant el desenvolupament neuronal, i que, quan està mutat, pot causar autisme. L’autisme és un trastorn que es caracteritza per afectar la conducta social, la capacitat d’atenció, la planificació i la comunicació de l’individu. Curiosament, altres gens relacionats amb l’autisme, com l’ACCN1 i el CADP2, i alguns d’associats a l’esquizofrènia (NRG3) i als trastorns cognitius de la síndrome de Down (DYRK1A), també semblen estar sota selecció positiva en humans moderns. Des de fa temps s’ha suggerit que l’espectacular desenvolupament cognitiu dels humans va comportar secundàriament l’aparició de nous trastorns mentals quan els gens implicats en aquest procés evolutiu pateixen mutacions. És a dir, la nostra malaltia mental és conseqüència de ser qui som, el que ara es diu, en contextos bèl·lics, un «dany col·lateral». Aquestes evidències semblen indicar que podria haver-hi diferències substancials entre nosaltres i els neandertals en aspectes cognitius que s’hauran d’investigar.
Altres gens situats en regions cromosòmiques amb rastres de selecció inclouen el gen THADA, que s’ha associat a diabetis de tipus II. Podria ser que aquest gen tingués una funció diferent en neandertals i humans moderns, i per tant, podria indicar també diferències en el metabolisme, potser associades a diferències en la dieta. Un altre gen interessant localitzat en una d’aquestes zones és el RUNX2. Sabem que, quan alguna mutació l’inactiva, es produeix un trastorn de l’ossificació (conegut com a displàsia clidocranial), que inclou el retard en el tancament de les sutures cranials, un os frontal anormalment bombat, una clavícula amb un desenvolupament incomplet i una caixa toràcica en forma de campana. Curiosament, alguns d’aquests trets exagerats pel trastorn corresponen a aspectes morfològics que difereixen entre nosaltres i els neandertals, els quals es caracteritzen, entre d’altres coses, per presentar cranis allargassats, arrodonits per darrere i amb el frontal poc vertical.
Tenim, doncs, evidències que alguna cosa passa amb gens com SPAG17, TTF1, RPTN, SOLH, TRPM1, NRG3, DYRK1A, RUNX2, THADA, AUTS2, CADP2 i ACCN1. A aquesta llista provisional de sospitosos s’haurien d’afegir altres dotzenes de gens que mostren algun canvi d’aminoàcid. Aquesta petita llista, que inclou gens implicats en la fisiologia de la pell, la pigmentació, la cognició, el desenvolupament ossi i el metabolisme, podria ser clau per definir la nostra espècie. Però per comprendre el que representen els canvis trobats, haurem d’estudiar-los un a un, i entendre’n la funció diferencial en humans moderns i en neandertals. La forma més fàcil d’aconseguir-ho serà la de crear ratolins transgènics amb aquests gens neandertals. La «neandertalització» de ratolins serà una de les conseqüències científiques del genoma neandertal, i implicarà força anys de recerques i possiblement de troballes espectaculars. Però ara per ara, estem començant a confeccionar la llista genètica on s’amaguen les claus de la nostra humanitat. No només sabrem com sentien, pensaven, menjaven i emmalaltien els neandertals, sinó que sabrem què és allò que ens fa diferents a nosaltres. Els neandertals, el nostre mirall evolutiu, ens ajudaran a definir-nos, 30.000 anys després de la seva extinció, i podrem ser, per primer cop, nosaltres mateixos.
Green, R. E. et al., 2010. «A Draft Sequence of the Neanderthal genome.» Science, 328 (5979): 710-722. doi: 10.1126/science.1188021




