

Biodiversitat marina en el temps i l’espai
Allò que ens diuen els fòssils minúsculs

La biodiversitat ha anat canviant tant en l’espai com en el temps. Per exemple, trobem més espècies en els tròpics i menys en la regió àrtica i l’antàrtica, seguint un gradient longitudinal de diversitat, un dels patrons que podem observar de manera més consistent en aquest món complex. El que sabem sobre els gradients temporals de biodiversitat és molt més limitat. Això ocorre perquè faria falta un programa de seguiment continu ben dissenyat, i aquests difícilment persisteixen més enllà d’unes dècades. Però, per sort, tenim restes d’antics organismes: els fòssils. Els fòssils són bàsicament l’únic registre directe de la biodiversitat passada.
Paraules clau: microfòssils, ecologia, evolució, paleontologia, biogeografia.
«Petits organismes coneguts com ‘microfòssils’ ens permeten inferir patrons de biodiversitat passats de forma detallada»
La bellesa i utilitat dels fòssils minúsculs
Alguns organismes tenen registres fòssils extraordinaris. En especial, els petits organismes coneguts com microfòssils (fòssils de grandària microscòpica) ens permeten inferir de manera detallada patrons de biodiversitat passats, ja que podem trobar-ne desenes de milers en un sol grapat de sediments i roques (Yasuhara, Tittensor, Hillebrand i Worm, 2017). Per exemple, coneixem el gradient longitudinal de diversitat de diversos organismes marins (Figura 1) (Hillebrand, 2004), però no és fàcil saber com era el gradient en el passat perquè, per exemple, els peixos (o la majoria de la resta d’organismes) rarament es conserven com a fòssils. Utilitzant un grup de microfòssils, els procedents de foraminífers planctònics (Figura 2), podem observar la biodiversitat del passat i el seu gradient longitudinal en el pèlag oceànic.
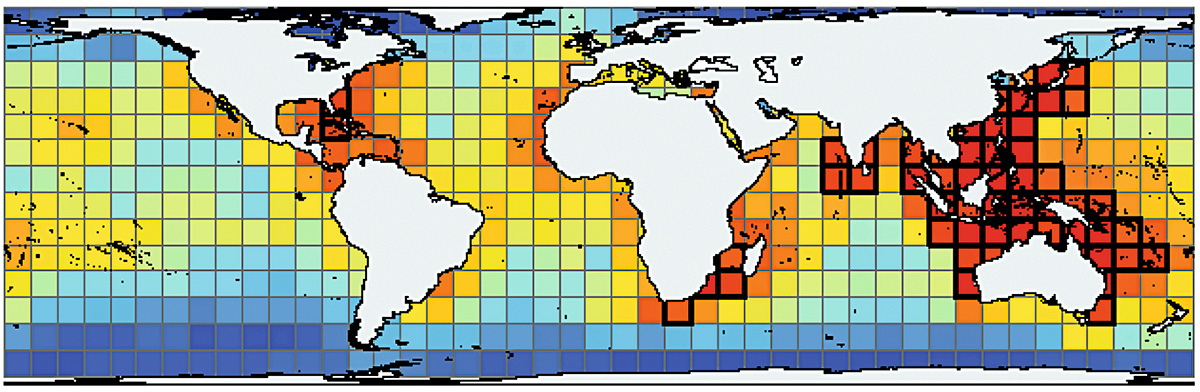
Figura 1. Patró mundial de biodiversitat marina. Els colors més càlids indiquen major biodiversitat (més espècies). / © Tittensor et al. (2010)
Potser els foraminífers planctònics (protozous amb closca que solen fer entre 0,1 i 1 mm) no són molt famosos, però són uns microfòssils molt importants per diverses raons. En primer lloc, se’n conserven milions en sediments i roques i representen un registre geològic i paleontològic quasi perfecte. En segon lloc, quasi totes les espècies cenozoiques s’han descrit formalment, la seua identificació estandarditzada està ben establerta per la seua importància en la bioestratigrafia i la paleoceanografía, i el nombre d’espècies és manejable (no són massa nombroses: se’n coneixen unes 40 espècies planctòniques en total en els oceans actuals); en altres paraules, la seua taxonomia és robusta (Yasuhara, Hunt, Dowsett, Robinson i Stoll, 2012; Yasuhara, Tittensor et al., 2017).
«Cada vegada és més important integrar les dades biològiques i els fòssils per a comprendre millor el nostre planeta»
Si comparem els patrons longitudinals de diversitat entre el present i l’última glaciació (fa 20.000 anys), observem la persistència del gradient que s’estén des dels tròpics fins als pols (Yasuhara et al., 2012) (Figura 3). Però analitzant els detalls és clar que el gradient de diversitat de la glaciació és molt més pronunciat que el present. Encara que les diversitats tropicals són semblants entre l’última glaciació i l’actualitat, les diversitats d’ambdós comencen a divergir a partir de 20–30º N i cap a latituds majors (Figura 3a). Per què? Si comparem les diversitats no sols amb la latitud sinó també amb la temperatura de l’oceà, trobem una possible resposta. Coneixem bé la temperatura superficial de l’oceà durant l’última glaciació gràcies a diferents mètodes micropaleontològics i geoquímics (Yasuhara et al., 2012). La relació entre temperatura i diversitat és sorprenentment consistent entre l’actualitat i l’última glaciació (Figura 3b). Per tant, el gradient longitudinal de diversitat era més pronunciat durant l’última glaciació perquè el fred sever en les latituds altes accentuava el gradient longitudinal de temperatura (Yasuhara et al., 2012).
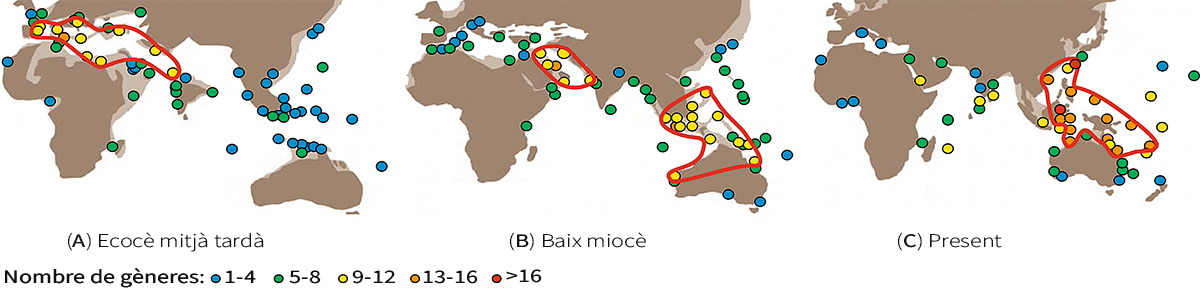
Figura 5. Distribució de la diversitat global dels gèneres de gran foraminífers bentònics (A) en l’eocè mitjà tardà (fa 42–39 milions d’anys), (B) en el baix miocè (fa 23–16 milions d’anys) i (C) en el present. Es pot observar que els punts calents de diversitat (assenyalats amb líneas roges) han anat variant. En l’eocè, la diversitat més alta es trobava en l’oceà de Tetis (l’antic Mediterrani), mentre que en el baix miocè, es trobava en la zona de la península d’Aràbia. El punt calent del triangle de corall de l’arxipèlag indoaustralià també va començar a formar-se en aquesta època, fa uns vint milions d’anys. Actualment, aquesta zona continua sent un punt calent de biodiversitat. / © Renema et al. (2008)
Punts calents de biodiversitat canviants
Fins i tot en els oceans tropicals, ben reconeguts com a regions biodiverses, la biodiversitat no roman estable en termes espacials ni temporals. Algunes àrees tenen moltes més espècies que altres. La diversitat d’espècies no varia només amb la latitud, com he explicat en la secció anterior, sinó també amb la longitud. El triangle de coral és un exemple d’això. Es caracteritza per la major biodiversitat marina del món (vegeu figures 1 i 4) (Bellwood, Renema i Rosen, 2012), però fins i tot la seua diversitat ha canviat amb el temps.
«Fins i tot en regions biodiverses com els oceans tropicals, la biodiversitat no roman estable en termes espacials ni temporals»
Els grans foraminífers (vegeu de nou la Figura 2), un tipus de foraminífers bentònics amb grans closques i algues simbiòtiques, mostren evidència directa que la biodiversitat del triangle de coral ha canviat. Potser no ens sorprendrà que la diversitat en aquesta àrea marítima s’haja mantingut persistentment alta en els últims vint milions d’anys, però, abans d’això, la biodiversitat de la regió era més modesta. En l’eocè, per exemple, la diversitat més alta es trobava en l’oceà de Tetis, l’antic Mediterrani (Figura 5) (Renema et al., 2008; Yasuhara, Tittensor et al., 2017).
«Els microfòssils ens ajuden a entendre millor els ecosistemes passats, presents i futurs»
El punt calent de biodiversitat del triangle de coral es va establir fa prop de vint milions d’anys, com també indiquen els fòssils i les dades moleculars d’altres organismes com els ostràcodes, els gasteròpodes i els peixos (Renema et al., 2008; Yasuhara, Iwatani et al., 2017). És probable que aquest origen del punt calent de biodiversitat del triangle de coral estiga relacionat amb la col·lisió d’Austràlia contra els arcs del Pacífic i el sud-est asiàtic fa entre vint-i-tres i vint-i-cinc milions d’anys, que va formar grans hàbitats marins superficials, diversos, complexos i dinàmics, la qual cosa va estimular la biodiversitat de la regió (Renema et al., 2008; Yasuhara, Iwatani et al., 2017; Yasuhara, Tittensor et al., 2017).
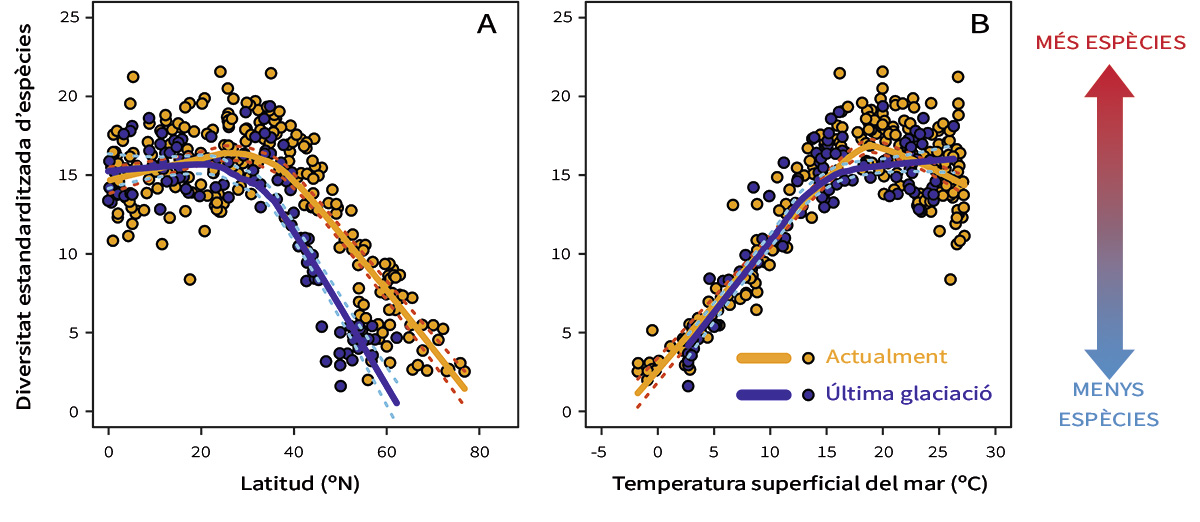
Figura 3. Gradient longitudinal de diversitat d’espècies per als foraminífers planctònics en l’actualitat i durant la darrera glaciació (fa 20.000 anys). Les línies són corbes suavitzades del diagrama de dispersió i ens permeten visualitzar una relació constant entre la temperatura de l’oceà i la biodiversitat de foraminífers planctònics, encara que el gradient de diversitat de la glaciació és més pronunciat que l’actual. Açò podria estar causat pel fred sever en les latituds altes durant l’última glaciació en comparació a la temperatura actual. Modificada a partir de Yasuhara et al. (2012).
Clima i biodiversitat d’aigües profundes
Les aigües profundes són un altre ecosistema caracteritzat per albergar nombroses espècies (Ingels et al., 2016; Rex i Etter, 2010). L’impacte climàtic en la biodiversitat d’aigües profundes és un tema que només es pot tractar de manera adequada si utilitzem els registres paleontològics, especialment els de microfòssils, perquè, en primer lloc, el nostre seguiment dels organismes marins moderns no sol ser prou prolongat per a poder avaluar els efectes dels canvis climàtics que ocorren en períodes de temps més llargs que la vida humana (Yasuhara, Doi, Wei, Danovaro i Myhre, 2016); i, segon, perquè la majoria dels organismes d’aigües profundes, especialment els grans, no tenen estructures mineralitzades (els bivalves i els gasteròpodes tenen closca però la seua abundància fòssil és massa baixa en els sediments d’aigües profundes) i, per tant, amb prou feines tenen registre fòssil.
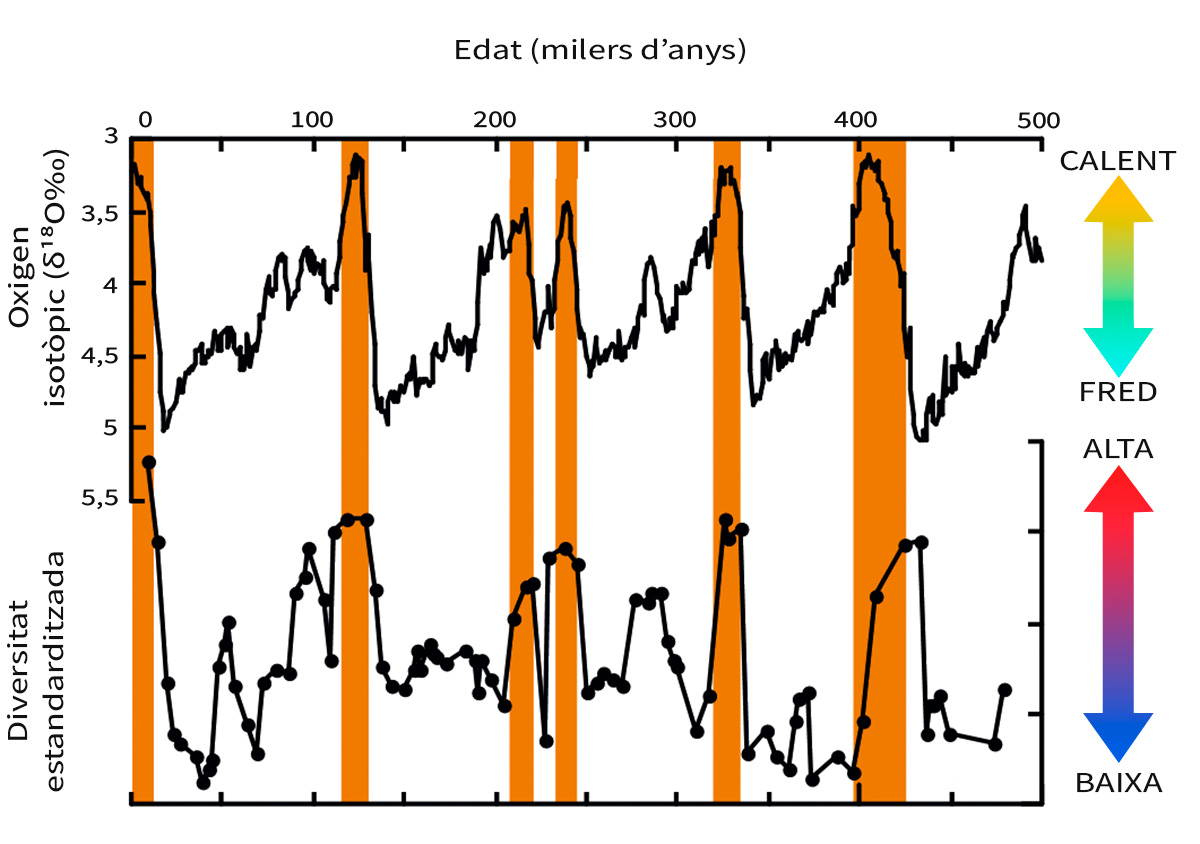
Figura 6. Comparació entre els canvis en el clima global i en la diversitat d’espècies d’ostràcodes d’aigües profundes durant els últims 500.000 anys. La corba isotòpica de l’oxigen (a dalt) representa els canvis climàtics globals, i els valors isotòpics més baixos indiquen intervals més càlids. Els pics principals de clima càlid estan marcats amb barres taronja. Es pot observar que la biodiversitat d’aquestes espècies d’ostràcodes és major en èpoques més càlides i menor durant les glaciacions. Modificada a partir de Yasuhara et al. (2009)
Els sediments dipositats en el fons de l’oceà són un arxiu ideal per a estudiar aquest tema perquè contenen nombrosos microfòssils, a més de registres climàtics excel·lents que es poden reconstruir mitjançant diversos mètodes geoquímics. Els microfòssils de bells crustacis petits (per exemple, ostràcodes com els que mostra la Figura 2, que en general fan aproximadament 1 mm o fins i tot menys i són molt sensibles als canvis ambientals, amb més de 20.000 espècies vives estimades) mostren una clara correlació amb els canvis climàtics glacials i interglacials (Cronin i Raymo, 1997; Yasuhara i Cronin, 2008; Yasuhara, Hunt, Cronin i Okahashi, 2009). La seua biodiversitat és major en èpoques més càlides i menor en les glaciacions (Figura 6) (Yasuhara et al., 2009). Coneixem bé aquest control tèrmic de la biodiversitat d’aigües profundes a diferents escales, des del present, fins a dècades o centenars de milers d’anys (Yasuhara i Danovaro, 2016).
Salvant les distàncies
Tradicionalment, les diferents comunitats d’investigació han estudiat la biologia i la paleontologia per separat. No obstant això, cada vegada és més important integrar les dades biològiques i els fòssils per a comprendre millor el nostre planeta i els organismes que hi viuen (Yasuhara, Tittensor et al., 2017). El nostre planeta està canviant ràpidament a causa de diferents activitats humanes, i conèixer les respostes passades a canvis semblants és molt important perquè podem preparar-nos millor per al futur. Gràcies al seu extraordinari registre fòssil, els microfòssils, encara que petits en grandària, tenen un paper crucial en la resposta a aquest problema.
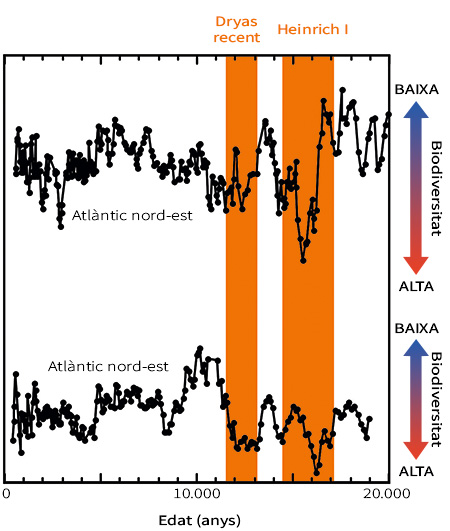
Figura 7. Canvis en la biodiversitat d’aigües profundes en el nord-est i el nord-oest de l’oceà Atlàntic durant els últims 20.000 anys. Els registres fòssils d’aquestes localitzacions tan distants (50.000 km) mostren augments de biodiversitat ràpids i sincrònics durant els successos de canvi climàtic ràpid del Dryas recent i el Heinrich I (marcats en taronja). Major biodiversitat significa més espècies en una mostra. Modificada a partir de Yasuhara et al. (2014)
Com a observació final, és notable que sapiguem tan poc sobre la manera com respon la biodiversitat marina als canvis climàtics ràpids en un entorn, al llarg de dècades o segles. Aquestes escales temporals són rellevants per als canvis antropogènics presents i futurs, com l’escalfament global, la desoxigenació, l’acidificació, la contaminació, l’augment del nivell del mar i la sobrepesca (Breitburg et al., 2018; Jackson et al., 2001; Mora et al., 2013). Aquestes escales de temps han estat un «punt cec» per als estudis biològics i paleoecològics, que són massa llargs per a l’observació humana i massa curts per a resoldre’s amb els registres fòssils ordinaris. Gràcies a registres de microfòssils excepcionalment detallats podem estudiar aquestes escales de temps. De fet, diferents estudis recents que han utilitzat registres de microfòssils han mostrat, per exemple, que la biodiversitat d’aigües profundes ha estat sensible a canvis climàtics abruptes a aquestes escales temporals (Yasuhara et al., 2016; Yasuhara, Cronin, DeMenocal, Okahashi i Linsley, 2008; Yasuhara, Okahashi, Cronin, Rasmussen i Hunt, 2014). Els registres de microfòssils mostraven evidents marques en la biodiversitat d’aigües profundes a causa de successos coneguts de canvi climàtic abrupte, com el Heinrich I (fa 17.000–14.600 anys) i el Dryas Recent (fa 12.900–11.700 anys) (Figura 7) (Yasuhara et al., 2014). Els models estadístics indicaven que aquests canvis en la diversitat van ser causats probablement per l’escalfament de les aigües profundes a nivell regional durant aquests successos de canvi climàtic abrupte (Yasuhara et al., 2014). Aquests canvis podrien haver afectat no sols la biodiversitat, sinó també el funcionament dels ecosistemes d’aigües profundes (Yasuhara et al., 2016). Els microfòssils ens ajuden a entendre millor els ecosistemes passats, presents i futurs, salvant la distància que separa l’escala temporal biològica i la paleontològica (Mesquita-Joanes, Smith i Viehberg, 2012; Yasuhara, Tittensor et al., 2017).
Referències
Bellwood, D. R., Renema, W., & Rosen, B. R. (2012). Biodiversity hotspots, evolution and coral reef biogeography: A review. En D. J. Gower, K. G. Johnson, J. E. Richardson, B. R. Rosen, L. Rüber, & S. T. Williams (Eds.), Biotic evolution and environmental change in Southeast Asia (pp. 216–245). Cambridge: Cambridge University Press.
Breitburg, D., Levin, L. A., Oschlies, A., Grégoire, M., Chavez, F. P., Conley, D. J., … Zhang, J. (2018). Declining oxygen in the global ocean and coastal waters. Science, 359(6371), eaam7240. doi: 10.1126/science.aam7240
Cronin, T. M., & Raymo, M. E. (1997). Orbital forcing of deep-sea benthic species diversity. Nature, 385, 624–627. doi: 10.1038/385624a0
Hillebrand, H. (2004). Strength, slope and variability of marine latitudinal gradients. Marine Ecology Progress Series, 273, 251–267. doi: 10.3354/meps273251
Ingels, J., Clark, M. R., Vecchione, M., Perez, J. A. A., Levin, L. A., Priede, I. G., … Van Gaever, S. (2016). Chapter 36F. Open Ocean Deep Sea. En L. Inniss & Simcock (Eds.), First global integrated marine assessment. World ocean assessment I. Nova York, NY: Organización de las Naciones Unidas.
Jackson, J. B. C., Kirby, M. X., Berger, W. H., Bjorndal, K. A., Botsford, L. W., Bourque, B. J., … Warner, R. R. (2001). Historical overfishing and the recent collapse of coastal ecosystems. Science, 293(5530), 629-638. doi: 10.1126/science.1059199
Mesquita-Joanes, F., Smith, A. J., & Viehberg, F. A. (2012). The ecology of Ostracoda across levels of biological organisation from individual to ecosystem: A review of recent developments and future potential. En D. J. Horne, J. Holmes, J. Rodriguez-Lazaro, & F. A. Viehberg (Eds.), Ostracoda as proxies for Quaternary climate change (pp. 15–35). Amsterdam: Elsevier.
Mora, C., Wei, C. L., Rollo, A., Amaro, T., Baco, A. R., Billett, D., ... Yasuhara, M. (2013). Biotic and human vulnerability to projected changes in ocean biogeochemistry over the 21st century. PLoS Biology, 11(10), e1001682. doi: 10.1371/journal.pbio.1001682
Renema, W., Bellwood, D. R., Braga, J. C., Bromfield, K., Hall, R., Johnson, K. G., … Pandolfi, J. M. (2008). Hopping hotspots: Global shifts in marine biodiversity. Science, 321(5889), 654–657. doi: 10.1126/science.1155674
Rex, M. A., & Etter, R. J. (2010). Deep-sea biodiversity: Pattern and scale. Cambridge: Harvard University Press.
Tittensor, D. P., Mora, C., Jetz, W., Lotze, H. K., Ricard, D., Berghe, E. V., & Worm, B. (2010). Global patterns and predictors of marine biodiversity across taxa. Nature, 466, 1098-1101. doi: 10.1038/nature09329
Yasuhara, M., & Cronin, T. M. (2008). Climatic influences on deep-sea ostracode (Crustacea) diversity for the last three million years. Ecology, 89(11), S53–S65. doi: 10.1890/07-1021.1
Yasuhara, M., Cronin, T. M., DeMenocal, P. B., Okahashi, H., & Linsley, B. K. (2008). Abrupt climate change and collapse of deep-sea ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 105(5), 1556–1560. doi: 10.1073/pnas.0705486105
Yasuhara, M., & Danovaro, R. (2016). Temperature impacts on deep-sea biodiversity. Biological Reviews, 91(2), 275–287. doi: 10.1111/brv.12169
Yasuhara, M., Doi, H., Wei, C. L., Danovaro, R., & Myhre, S. E. (2016). Biodiversity-ecosystem functioning relationships in long-term time series and palaeoecological records: Deep sea as a test bed. Philosophical Transactions of the Royal Society B, 371(1694). doi: 10.1098/rstb.2015.0282
Yasuhara, M., Hunt, G., Cronin, T. M., & Okahashi, H. (2009). Temporal latitudinal-gradient dynamics and tropical instability of deep-sea species diversity. Proceedings of the National Academy of Sciences of the United States of America, 106(51), 21717–21720. doi: 10.1073/pnas.0910935106
Yasuhara, M., Hunt, G., Dowsett, H. J., Robinson, M. M., & Stoll, D. K. (2012). Latitudinal species diversity gradient of marine zooplankton for the last three million years. Ecology Letters, 15(10), 1174–1179. doi: 10.1111/j.1461-0248.2012.01828.x
Yasuhara, M., Iwatani, H., Hunt, G., Okahashi, H., Kase, T., Hayashi, H., … Renema, W. (2017). Cenozoic dynamics of shallow-marine biodiversity in the Western Pacific. Journal of Biogeography, 44(3), 567–578. doi: 10.1111/jbi.12880
Yasuhara, M., Okahashi, H., Cronin, T. M., Rasmussen, T. L., & Hunt, G. (2014). Response of deep-sea biodiversity to abrupt deglacial and Holocene climate changes in the North Atlantic Ocean. Global Ecology and Biogeography, 23(9), 957–967. doi: 10.1111/geb.12178
Yasuhara, M., Tittensor, D. P., Hillebrand, H., & Worm, B. (2017). Combining marine macroecology and palaeoecology in understanding biodiversity: Microfossils as a model. Biological Reviews, 92(1), 199–215. doi: 10.1111/brv.12223




