|
In times of the great dinosaurs. The giant Sauropod that lived in the Valencia region 130 million years ago. The first remains of giant herbivorous Dinosaurs, quadrupeds, that we call Sauropods, became known in 1841 thanks to the discoveries made by the Englishman OWEN. From this moment on, the number of remains found has increased, but it was not until the beginning of the XX Century when the first remains from the Jurassic and Cretaceous periods in Spain, specifically in the eastern region. However, in the second half of this century, due to the first excavation works, we have more and more remains at our disposal, although, mostly, they are unconnected and difficult to interpret. Despite this, since 1987 three taxa, new to science, have been found, up to now exclusively Spanish: Aragosaurus ischiaticus in the site at Las Zabacheras (Galve, Teruel); Lirainosaurus astibiae in Laño (Condado de Treviño, Burgos) and Losillasaurus giganteus in Losilla (Comarca de Los Serranos, Valencia). Possibly to these we can add a new taxon after studying the remains found in Peñarroya de Tastavins (Teruel). Els sauròpodes: gegants herbívors Els dinosaures sauròpodes eren gegants herbívors, quadrúpedes que es reproduïen mitjançant ous. Van aparèixer a començament del Juràssic i van sobreviure fins al Cretaci final, essent els herbívors terrestres dominants durant la major part de l’esmentat període. El coneixement que en tenim comença el 1841, a Anglaterra, quan Owen descriu la primera resta trobada i li dóna el nom de Cetiosaurus. S’hi afegeixen després noves restes que es van assignar a aquest sauròpode (Mantell, 1850; Gervais, 1852), però fins a 1871 aquests animals no van començar a ser coneguts, gràcies a la descripció que Phillips va fer de gran part d’un nou esquelet atribuït a Cetiosarus oxiniensis, del Juràssic Mitjà d’Anglaterra. Durant les dècades del 1870 a 1890 autors com ara els anglesos Seeley i Hulke van trobar solucions a la major part dels problemes referents a l’anatomia dels sauròpodes, però els coneixements van augmentar espectacularment el 1877, quan va tenir lloc la gran explosió de descobriments amb esquelets moltes vegades articulats a l’oest d’Amèrica del Nord (Colorado i Wyoming, Juràssic Superior, formació Morrison), deguts principalment als professors Cope i Marsh. De 1890 a 1910 s’hi van unir els primers descobriments de sauròpodes a Sud-amèrica (Argentina) i Àfrica (Madagascar). Des de llavors van anar eixamplant-se les localitats i països on s’han trobat restes de sauròpodes i han continuat les exploracions en tot el món. Especialment interessants han estat els avanços dels estudis a Xina i Mongòlia; i espectaculars els resultats obtinguts, de manera que, actualment, podem dir que s’han trobat sauròpodes en tots els continents excepte a l’Antàrtida. A principis del segle XX alguns paleontòlegs espanyols, en especial el professor Royo i Gómez, va explorar el Llevant espanyol (voltants de Morella en particular), amb resultats que no van poder arribar a publicar. És a partir del segon terç del segle XX quan s’obtenen resultats positius que han estat estudiats i publicats i que demostren que també a la Península Ibèrica van viure els grans dinosaures herbívors que ens ocupen i que la majoria de les vegades presenten característiques particulars que els allunyen dels anteriorment descrits i que mereixen que se’ls dedique un apartat especial. Consideracions paleoecològiques Les formacions en què s’han trobat la major part de les restes fan pensar que l’hàbitat principal dels sauròpodes, encara que terrestre, era de caràcter humit, i que vivien prop d’ambients aquàtics dolços o salobres. Concentracions d’aquestes restes es van trobar recobertes de petxines marines (jaciments del Canteret, Morella), cosa que va fer suposar als autors la proximitat del mar en el Cretaci Inferior, encara que la línia de costa no estava connectada directament amb el mar obert perquè hi havia un estany limitat per un complex d’“illes barreres”. S’han trobat abundants empremtes al costat de llacunes o restes d’antics mars que s’anaven retirant, la qual cosa confirma que els sauròpodes realment caminaven sobre estrats coberts per aigües somes o recentment submergides. En canvi a Amèrica del Nord, en la formació Morrison, les faunes de sauròpodes són essencialment homogènies en distàncies de 1.000 km. Aquesta distribució, afegida a les proves proporcionades per l’estudi sedimentològic de l’existència d’estacions àrides així com de l’absència de restes de plantes fa suposar que els sauròpodes devien emigrar recorrent grans distàncies a la recerca d’aliments com fan actualment a Àfrica els moderns ungulats. Hem de tenir en compte que a principis del mesozoic encara hi havia un únic continent, de manera que no és estrany que s’hagen trobat sauròpodes del mateix gènere en cares oposades de continents juxtaposats. Les seues llargues extremitats indubtablement els ajudaven en els seus extensos recorreguts i segons alguns autors, els permetien fer 10 o 20 quilòmetres per dia. A través de les pistes deixades durant el seu trajecte s’ha pogut detectar un suposat gregarisme en alguns gèneres, encara que realment no es pot assegurar que els sauròpodes foren necessàriament gregaris. També s’ha especulat sobre l’ús o no de gastrolits per a ajudar-se en la digestió. La discussió se centra en la simplicitat de les dents –molt dèbils per a la masticació– i en la presència de pedres polides de contorn arrodonit i proveïdes d’una pàtina característica, trobades entre la pelvis i les costelles d’algun espècimen. Un altre punt àmpliament discutit per diversos autors és la possibilitat que foren endotèrmics. Des de la primera hipòtesi fins a l’actualitat, defensors i detractors han entaulat batalles científiques i fins al moment no sembla que s’haja arribat a cap conclusió definitiva. Resum diagnòstic: gran cos i crani petit La diagnosi dels sauròpodes es basa en la gran talla del seu cos, els petits cranis i els llargs colls i cues. El crani té els orificis nasals situats dorsalment i, la major part de les vegades, l’os jugal està exclòs de la vora ventral del crani. La capacitat cranial era relativament petita i les dents molt simples. Els centres presacrals de les vèrtebres presenten pleurocels fortament enfonsats, amb arcs neurals àmpliament reduïts a un complex de primes làmines. Solen tenir de 12 a 19 vèrtebres cervicals i de 8 a 14 dorsals. L’escàpula està orientada més horitzontalment que verticalment; l’ili té el procés acetabular àmpliament expandit i el peduncle púbic més llarg que l’isquiàtic. Els ossos de les extremitats són robustos i sòlids, sense entalladura entre el cap i el trocànter major en el fèmur. El carp i el tars estan reduïts a un o dos elements cada un i en les formes més primitives algunes vegades els metacarpals són més llargs que els metatarsals. El nombre de falanges de les mans està reduït, de manera que el primer dígit només té una ungla. També estan reduïdes el nombre de falanges en els dits quart i cinquè del peu. Història de la classificació Les idees referents a la classificació de sauròpodes han anat canviant al llarg dels anys i continua sent així. L’esquema multifamiliar de Marsh va prevaler fins a 1929, quan Janensch va proposar un esquema de dues famílies basat en la forma de les seues dents. Aquells que tenien les dents espatulades van ser situats en els Brachiosauridæ, enfront d’aquells que les tenien en forma d’estaca, que va col·locar en els Titanosauridæ. El resultat d’aquest esquema va ser incloure en la mateixa família els diplidòcids i els titanosàurids, dos grups de sauròpodes completament divergents els uns dels altres. Només tenien un caràcter comú: les dents. El 1986, trobem intents de classificació basats ja en l’estudi de sinapomorfies que constitueixen el punt de partida per a establir una diagnosi cladística. En el mateix any, Bonaparte fa un estudi comparatiu de les vèrtebres sacrals dels sauròpodes en què posa especial atenció en caràcters avançats presents en la majoria d’aquests però absents en les formes basals. La presència de pleurocels i la marcada opistocèlia a les vèrtebres presacrals formaven la base de la distinció de dos graus: eusauròpodes (Juràssic Mitjà-Final), i neosauròpodes per als del juràssic terminal. És curiós que no incloguera els titanosauris en la seua definició de neosauròpodes, com digueren recentment Wilson i Sereno, perquè deixa incompleta la classificació. McIntosh proposa una nova classificació a finals de la dècada de 1980, basada en una acurada consideració de disset regions variables en l’esquelet. Com a conseqüència presenta un arbre filogenètic que té Vulcanodon com a base dels següents sauròpodes. Des de llavors nous descobriments i estudis recents han donat lloc a distintes classificacions. Poc desprès, Upchurch presenta el treball més consistent de la relació filogenètica dels sauròpodes realitzat fins al moment, basat en l’anàlisi cladística de 174 caràcters en 27 gèneres, tot i que la seua avaluació de les hipòtesis anteriors i el tractament d’aquests caràcters, així com les seues conclusions, són també àmpliament discutits per Wilson i Sereno. Aquests autors arriben a la conclusió que, encara que Upchurch indica diverses sinapomorfies de clades de sauròpodes basals, la seua cladística, fundada en la tradicional associació de diplodòcids i titanosàurids, és dèbil i basada principalment en caràcters dentals ambigus i sustentada pobrament en caràcters postcranials. En la dècada de 1990, Calvo i Salgado van presentar una anàlisi filogenètica i, junt amb Rodolfo Coria, fan un nou estudi treballant especialment els titanosàurids i la relació d’aquests amb els altres grups. La contribució més important en aquest estudi és la proposta d’usar el terme titanosauriformes, com a nou clade que inclou Brachiosaurus i Titanosauria. Dinosaures sauròpodes a Espanya Els primers treballs d’excavació sensu stricto realitzats a Espanya es van realitzar pels volts de Morella. Un veí d’aquesta ciutat, Francisco Yeste, va descobrir el jaciment del Canteret (Cretaci Inferior) que va proporcionar abundants restes d’un sauròpode que es va preparar i estudiar en la mateixa Morella. En la publicació de les restes trobades en aquest lloc (1982), els autors assignen aquest sauròpode a la família Brachiosauridæ de Riggs 1904, en el sentit que McIntosh donava a aquesta i d’acord amb la relació entre les longituds de l’húmer i el fèmur, ja que una de les característiques que s’atribuïen a aquestes famílies eren la major longitud de l’extremitat anterior respecte a la posterior. En les restes del Canteret s’han tingut en compte, doncs, les dimensions longitudinals de l’húmer i el fèmur; la gràcil morfologia humeral i l’aparició d’un tubercle femoral característic, a la zona proximal externa de la diàfisi. Les restes estudiades, però, no van permetre fer una assignació més precisa. Actualment José Miguel Gasulla està fent una revisió dels dinosaures morellans incloent-hi el nou material que s’ha recuperat des de la primera publicació i possiblement podrà arribar a establir una conclusió més ferma quant a la seua assignació. La segona excavació va tenir lloc als voltants de Galve, província de Terol. El 1960 el professor Lapparent havia descrit alguna resta de sauròpode no identificat del jaciment de Las Zabacheras (Cretaci Inferior). No obstant això, posteriorment, amb el material procedent de la col·lecció de José María Herrero, veí de Galve, es van identificar quatre formes diferents de sauròpodes, si bé tres d’aquestes només es van definir mitjançant peces dentàries aïllades, el que posa en dubte la seua assignació. La quarta forma, el sauròpode de Las Zabacheras (Hauterivià Superior), és representada per les restes d’un únic individu amb característiques molt especials que va donar peu a la creació d’un nou taxó amb la denominació d’Aragosaurus ischiaticus (“saure isquiàtic d’Aragó”). El terme ischiaticus fa referència a l’especial desenvolupament dorsoventral de l’apòfisi púbica de l’isqui, un element que sembla especialment diagnòstic entre els sauròpodes. Els autors esmentats inclouen aquest nou sauròpode entre els camarasàurids, però, davant els darrers avenços de la classificació supraespecífica dels sauròpodes, José Ignacio Canudo analitza de nou gran part del material d’Aragosaurus ischiaticus, que, fins el 1997, era encara l’únic gènere del Cretaci Inferior inclòs en la família adés esmentada. Segons aquests autors “la presència d’un bombament lateral ben desenvolupat en el fèmur d’Aragosaurus” indica que es troba situat en el clade dels titanosauriformes i que no es tracta, doncs, d’un representant de la família Camarasauridae. Els titanosauriformes constitueixen un clade de sauròpodes àmpliament distribuït arreu del món. Fan la seua aparició en el registre fòssil en el Juràssic Superior i s’estenen fins al Cretaci final amb els pròpiament denominats titanosauris. Seguint l’ordre de descobriments de nous sauròpodes a Espanya podem citar Lirainosaurus astibiae (Sanz, Powell, Le Loeuff, Martínez i Pereda, 1999) (“saure gràcil d’Astíbia”), d’acord amb les restes trobades al jaciment de Laño (Cretaci Superior), situat al comtat de Treviño (centre nord d’Espanya). El terme lirain és un mot basc que significa “prim”. El nou taxó deu pertànyer als titanosauris. Fins fa poc temps es pensava que els titanosauris eren dinosaures típicament gondwanians, alguns dels quals havien envaït els continents del nord durant el Cretaci Superior després del tancament del Tetis. Troballes recents, però, podrien modificar aquests supòsits. Com a últim taxó, específicament hispà i llevantí, trobat fins ara, hem de citar Losillasaurus giganteus, situat en el trànsit Juràssic Superior – Cretaci Inferior i per això és, fins ara, el sauròpode més antic d’Espanya. Després de trobar in situ (1989) un os gran a la localitat de Losilla (Serrans), a l’estiu del mateix any es va començar l’excavació sistemàtica d’aquest jaciment que es va prolongar fins a 1995. Les restes trobades són de grans dimensions i estaven embolicades en una matriu extremadament dura que en dificultava l’extracció. També va ser molt laboriosa la preparació del material i el seu estudi no es va poder realitzar fins que se n’obtingueren rèpliques en resina polièster prou manejables. A partir de 1996 es va obtenir nou material de sauròpode en Pena-roja de Tastavins (Matarranya). Es tracta d’un esquelet parcialment articulat d’un únic individu, possiblement un dels sauròpodes més complets del món que es coneixen del Cretaci Inferior. Actualment està en estudi i encara no en tenim una definició científica. El llangraix gegant de Losilla: Losillasaurus giganteus Figura 4. Reconstrucció del jaciment de dinosaures de Losilla (València). Els “ossos”de Losillasaurus giganteus són reproduccions dels originals, que es troben al Museu de Ciències Naturals de València. Losillasaurus giganteus (“llangardaix gegant de Losilla”) és el tercer dinosaure sauròpode d’origen espanyol, és a dir, que les seues restes han estat descobertes i estudiades per primera vegada al nostre país. El material fòssil amb què es va proposar l’any 2001 el nom de Losillasaurus consta de més de trenta fragments i elements esquelètics complets. Aquesta evidència inclou fragments cranials, vèrtebres del coll, del tronc i de la cua, ossos de l’extremitat anterior, de l’estèrnum i de la cintura pèlvica (malucs). El lector es deu preguntar si amb tan exigu material (que representa un percentatge mínim del nombre total d’ossos que l’animal devia tenir) es pot saber que es tracta d’una espècie (i gènere) nous per a la ciència. La resposta és sí. Aquesta contundent afirmació es basa en el fet que determinats elements esquelètics de Losillasaurus presenten diferències en morfologia que eren completament desconegudes, fins llavors, en la resta dels dinosaures registrats. Aquesta singularitat es localitza a les vèrtebres anteriors de la cua. En tots els dinosaures la projecció dorsal (neuroapòfisi) d’aquestes vèrtebres és recta, mentre que en Losillasaurus gira cap arrere, en forma d’“alfange”. A més, la seua base té una dimensió anteroposterior molt més desenvolupada que en la resta dels dinosaures. Aquestes característiques ens van permetre saber que el material de Losilla pertanyia a una forma nova, però no ens en facilitaven la identificació perquè no les comparteix amb cap altre dinosaure conegut. Per tant, hem de fixar-nos en altres trets de l’esquelet que compartesca amb algun llinatge de dinosaures sauròpodes. El tret principal que cal tenir en compte és la morfologia de les projeccions laterals de les vèrtebres caudals anteriors, en forma d’ala. Aquestes estructures aliformes apareixen en dinosaures com Diplodocus i les seues formes emparentades (com per exemple Barosaurus i Dicraeosaurus). Per tant, creiem que Losillasaurus devia ser un parent pròxim d’aquest grup, encara que més primitiu, ja que manca d’altres novetats evolutives que diagnostiquen al gènere Diplodocus i les seues formes afins. Losillasaurus giganteus va habitar el que avui dia és la comarca dels Serrans durant el trànsit entre el Juràssic Superior i el Cretaci Inferior, fa uns 140-130 milions d’anys del present. El seu húmer fa una longitud de 143 cm, la qual cosa indica que l’animal en vida devia tenir més de 20 metres de longitud. Es tracta d’un enorme dinosaure menjador de plantes, encara que desconeixem quines en concret. Maria Lourdes Casanovas Cladelles. Institut de Paleontologia “Miquel Crussafont”, Sabadell. |
Figura 1: Motlle de l’húmer esquerre complet de Losillasaurus giganteus. En aquesta fotografia es pot apreciar la grandària que va tenir en vida aquest sauròpode. En la fotografia el professor José Luis Sanz.
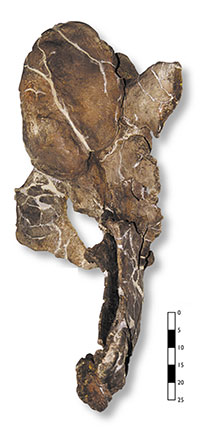
Figura 2. Vista posterior d’una vèrtebra caudal de posició anterior. Observeu la forma alar de les apòfisis transverses.
Figura 3. La mateixa vèrtebra caudal, en vista lateral dreta, mostra la característica més significativa d’aquest gènere, que correspon a la forma d’alfange de la neuroapòfisi.
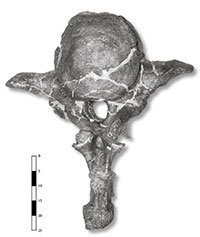
Figura 4. Vèrtebra caudal de posició mitjana-anterior, en vista posterior. Al llarg de la cua, es perd la morfologia alar de l’apòfisi transversa i es verticalitza la neuroapòfisi. |