

Gens arquitectes del cervell
L’evolució cerebral a través de la genoarquitectura
El cervell dels humans moderns és el resultat de l’evolució d’un pla de construcció (Bauplan) que comença a dissenyar-se fa 500 milions d’anys. Va ser un procés iniciat en els cordats basals (animals marins que vivien immersos en l’arena) que va donar origen al primer pla de construcció del sistema nerviós central; aquest es va modificar progressivament i el van compartir tots els vertebrats. Darrere d’aquest procés hi ha les xarxes gèniques, actors clau per a donar identitat a les distintes regions del cervell. Aquest escenari evolutiu proporciona la base per als estudis que tracten d’entendre el que es «conserva» i el que és «nou» entre distints vertebrats, així com també dels mecanismes subjacents implicats en aquest procés. Aquest article explora el paper dels estudis genoarquitectònics en aquesta aventura científica.
Paraules clau: evolució del cervell, patrons d’expressió gènica, regionalització cerebral, xarxes gèniques.
És el cervell humà diferent del d’un tauró, un pollastre o un ximpanzé?
Tots els vertebrats tenen un origen evolutiu comú, és a dir, que tenen un avantpassat comú. Un dels principals passos en el llarg camí per a produir el cervell dels vertebrats actuals era obtenir un pla de disseny bàsic per a la construcció del cervell. Una manera de comprendre el significat d’aquest pla de construcció és comparar-lo amb els plànols d’una casa. Des del començament de l’evolució cultural de la humanitat, el model de casa s’ha modificat des d’un únic compartiment per a usos múltiples, fins al model actual, que té distintes «parts», com la cuina, el dormitori, el bany i la sala d’estar. Aquestes «parts» es van anar agregant segons necessitats pròpies de cada època; per exemple, un aparcament en etapes més recents. No obstant això, totes les cases tenen els mateixos espais bàsics. Un fenomen semblant va ocórrer amb l’evolució del cervell dels vertebrats. Els cordats basals de fa 500 milions d’anys van donar origen a un sistema nerviós central relativament simple amb pocs compartiments; aquest model va ser modificat quan van sorgir els vertebrats i va portar al desenvolupament d’un nombre més gran de regions.
«Com sabem si una estructura anatòmica és diferent d’una altra? Un fet que pareix senzill, però que no resulta una tasca fàcil»
Diversos estudis experimentals han demostrat que el cervell de tots els vertebrats parteix d’un pla general que s’estableix ben aviat durant el desenvolupament. La primera cosa que ocorre durant la formació del sistema nerviós central és la generació d’un tub neural a partir d’una làmina de cèl·lules coneguda com a placa neural. Aquest procés s’esdevé en l’ésser humà durant la tercera i quarta setmana de desenvolupament embrionari. A partir d’aquest moment la placa i el tub comencen a «regionalitzar-se» tant en la seua dimensió rostrocaudal (anteroposterior) com en la dorsoventral (és a dir, entorn de la circumferència del tub) (Nieuwenhuys, Voogd i Van Huijzen, 2008). Així es produiran des de la part més rostral a la caudal del tub neural les diferents regions que constitueixen el sistema nerviós central. La part més rostral del tub produeix el cervell anterior o prosencèfal, que dóna lloc a l’hipotàlem i al telencèfal (que inclou l’escorça cerebral) en la seua part rostral, i al diencèfal caudalment (que produeix el pretàlem, tàlem i presostre). Caudalment al prosencèfal trobem el cervell mitjà o mesencèfal, el cervell posterior o romboencèfal (que dóna lloc al cerebel entre altres estructures) i la medul·la espinal (figures 1 i 2).
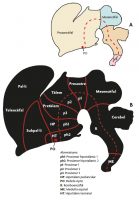
Avui dia sabem que aquestes regions contenen compartiments anteroposteriors més petits que funcionen com a blocs de construcció, coneguts com neuròmers; per exemple, el prosencèfal inclou cinc neuròmers: dos més de rostrals que donen lloc a l’hipotàlem i al telencèfal; i tres més de caudals que donen lloc a distintes parts del diencèfal (figura 1). Al seu torn, cada neuròmer se subdivideix en compartiments més petits al llarg de l’eix dorsoventral. Els detalls de la manera com es regionalitza el cervell de vertebrats s’aborden amb precisió en el model prosomèric proposat per Puelles i Rubenstein (2003, 2015). Segons aquest model, aquests compartiments fonamentals es troben en tots els vertebrats i s’estableixen durant el seu desenvolupament embrionari primerenc; per tant, representen les unitats de desenvolupament que formen part del pla de construcció del sistema nerviós central dels vertebrats. Això significa que trobarem els mateixos compartiments generals (els mateixos neuròmers i les seues subdivisions dorsoventrals) tant en cocodrils, ànecs, elefants com en humans.
Ara bé, si els compartiments són els mateixos, en què difereixen aquests cervells entre si? La principal diferència són els derivats que cadascun pot donar. Un dels exemples més interessants correspon a l’escorça cerebral, una part del telencèfal molt expandida en molts mamífers (figura 2), però amb una organització molt diferent en no-mamífers. Això significa que un determinat compartiment pot produir estructures diferents en grandària i morfologia en aus i mamífers. De fet, actualment encara es discuteix exactament quina és la regió comparable (homòloga) a l’escorça cerebral de mamífers en aus i rèptils. D’altra banda, quan es compara l’escorça cerebral de diferents mamífers, com ara rosegadors, gats, ximpanzés i humans, es fan evidents les diferències de grandària, molt destacable en primats i, en particular, en humans.
En conclusió, si bé tots els vertebrats tenim el mateix pla de construcció del cervell, cada espècie ha pogut variar la grandària de les seues divisions i fins i tot produir «nous» derivats. Reprenent l’exemple de les parts bàsiques d’una casa: tots els habitatges tenen cuina però aquesta no és igual en totes les llars, perquè no totes les famílies tenen les mateixes necessitats; el mateix s’aplica al cervell comparat entre els distints vertebrats. Les mateixes parts són presents en tots, però pot canviar la grandària, el nombre o el tipus de derivat que trobem en cada regió, o fins i tot la localització (si hi ha hagut alguna variació en la migració de les neurones produïdes en cada compartiment durant el desenvolupament). La pregunta que en aquest punt hauríem de fer-nos és: com s’ha arribat a aquestes conclusions? O dit amb altres paraules, quines són les estratègies d’investigació que permeten acostar-nos a aquesta idea? Ací és on els gens comencen a prendre un rol protagonista, i estudiant-los els investigadors actuen com detectius per a intentar conèixer què pot haver succeït durant l’evolució del cervell.
Quin és el paper dels gens en el desenvolupament del cervell?
Els gens, quan estan actius, sintetitzen molècules d’ARN (mitjançant un procés conegut com a transcripció) que després s’utilitzen per a produir una proteïna (a través del mecanisme denominat traducció). Aquesta última molècula és la que finalment exerceix distintes funcions, com les relacionades amb el manteniment de la cèl·lula, l’especificació de la identitat cel·lular durant el desenvolupament, l’augment del nombre de cèl·lules (proliferació), els canvis en l’adhesivitat, etc. L’activitat d’un gen és controlada per regions d’ADN la funció de les quals és regular l’expressió gènica. Aquestes zones reguladores determinaran quan i on s’expressarà un gen, activant o reprimint les regions promotores que donen inici a la transcripció. Alguns gens tenen com a funció primordial regular l’expressió d’altres. Els seus productes proteics, que es coneixen com a factors de transcripció, es transporten al nucli i interactuen amb les regions reguladores que controlen l’expressió d’altres gens. L’activació d’un gen sol requerir d’un grup de productes proteics d’altres gens; a més, en alguns casos, la seua pròpia activació pot conduir a activar o reprimir l’expressió d’altres gens. Encara que presentat ací de manera simplificada, aquest model de funcionament en què diversos productes gènics (proteïnes) poden interactuar determinant l’expressió d’altres gens, és subjacent a la manera com funciona una xarxa gènica (Davidson, 2006; Puelles i Ferran, 2012).
«La genoarquitectura neural fa referència a la descripció de les estructures neurals en termes de patrons d’expressió gènica, i implica l’ús de les sondes de ARNm com a marcadors morfològics»
Durant el desenvolupament del cervell, el tub neural va dividint-se en compartiments, que adquireixen una identitat pròpia a causa de la participació de xarxes gèniques que s’activen de manera diferencial en l’espai (i que donen lloc a distintes regions) i en el temps (en cada moment del desenvolupament). Els derivats anatòmics que s’originen progressivament a partir de cada compartiment estaran estretament relacionats amb els efectes d’aquestes xarxes gèniques. Ara bé, el nombre de gens que posseïm els humans no és infinit i voreja els 20.000. La major part són utilitzats en el cervell, ja siga per construir-lo o per fer-lo funcionar. Considerant la gran diversitat de processos de regionalització i especificació des que comença a formar-se l’organisme i la gran varietat de tipus cel·lulars que posseeix cada òrgan, aquest nombre de gens és massa baix perquè es puga associar un gen a una funció. Un producte gènic sol utilitzar-se tant en el desenvolupament com en la vida postnatal en multiplicitat d’esdeveniments. És el context en què s’expressen, és a dir, amb quins altres gens interactuen i la història molecular prèvia de la regió, allò que determinarà l’efecte final en l’òrgan. Per tant, la qüestió important en aquest procés són les xarxes d’expressió de gens.
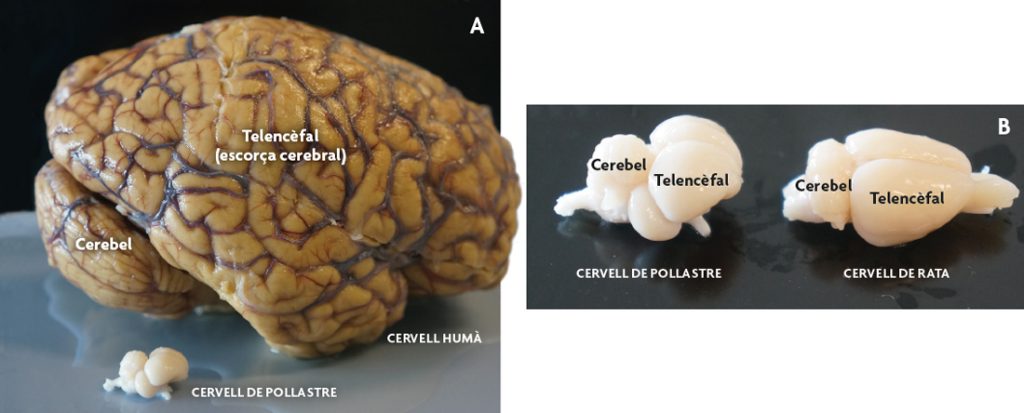
Figura 2. Sistema nerviós central de mamífers i aus. A) Fotografia comparant un cervell humà amb un de pollastre. Cal destacar la grandària de l’escorça cerebral humana. B) Vista d’un cervell de pollastre d’un mes de vida postnatal i d’un cervell de rata de dos mesos i mig de vida postnatal (adult jove). En el rosegador l’escorça cerebral també està expandida, però a diferència de l’humà no presenta plecs. No obstant això, en el pollastre el telencèfal correspon majoritàriament a estructures diferents de l’escorça cerebral (actualment encara es discuteix quina és la regió homòloga a l’escorça cerebral en aus). En tots els casos es destaca el cerebel (part del cervell posterior o romboencèfal). / José Luis Ferran
Un dels esdeveniments clau en l’evolució dels vertebrats i específicament del sistema nerviós central va ser una doble ronda de duplicació del genoma sencer que va ocórrer en els avantpassats dels vertebrats. Susumu Ohno (1970) va proposar la denominada teoria de la doble duplicació genòmica (hipòtesi 2R) com a esdeveniment clau en l’origen dels vertebrats. Aquesta proposta ha estat avalada pels resultats de l’anàlisi de seqüències genòmiques completes de diferents vertebrats i de cordats basals com el de l’actual amfiox (Putman et al., 2008). Com a conseqüència d’aquestes duplicacions, els vertebrats podrien tenir potencialment quatre còpies de cada gen. Aquesta redundància d’informació va permetre d’una banda que es conservaren funcions inicials, i d’una altra que se’n generaren de noves o se separaren funcions entre els «nous» gens. Encara que s’han perdut molts gens producte d’aquella duplicació com a resultat del procés evolutiu, actualment el genoma d’aus i de mamífers té més d’una còpia per a molts dels seus gens. Alguns peixos teleostis i espècies d’amfibis del gènere Xenopus tenen rondes addicionals de duplicacions genòmiques. La doble duplicació genòmica no va ser l’únic mecanisme que va incrementar el nombre de gens durant aquests 500 milions d’anys, sinó que a més en molts casos els gens es van duplicar individualment i es van alinear l’un darrere de l’altre. L’augment del nombre de gens va anar paral·lel a l’augment de complexitat dels organismes i en el cas del sistema nerviós central podem trobar un pla de construcció més complex en vertebrats que l’observat en cordats basals.
«La genoarquitectura permet obtenir detalls sobre l’origen durant el desenvolupament i l’evolució dels distints grups de neurones del cervell no imaginables dècades arrere»
Els gens que treballen en xarxa i en localitzacions anatòmiques específiques estan implicats en distintes funcions depenent del context. Si el pla de formació del sistema nerviós central es conserva o es comparteix entre vertebrats, esperaríem trobar xarxes gèniques molt semblants funcionant en la mateixa regió del cervell de distints vertebrats. No obstant això, també cal considerar que, en alguns casos, en una determinada localització aquestes xarxes podrien haver canviat de manera significativa durant l’evolució. Com analitzarem en el pròxim apartat, els productes gènics que actuen en etapes primerenques del desenvolupament i participen en l’especificació de la identitat d’una regió o dels seus derivats són d’utilitat per a estudis comparats, perquè permeten reconèixer quins components s’originen a partir de la mateixa regió en les distintes espècies.
La genoarquitectura ajuda a entendre l’evolució del cervell
Com sabem si una estructura anatòmica és diferent d’una altra? Un fet que sembla senzill, però que no és una tasca fàcil. Si el que volem és saber com reconèixer una estructura anatòmica i destriar-la d’una altra, necessitem eines potents que indiquen clarament l’extensió d’un territori (els seus límits o vores), o la identitat comuna que pot tenir un grup de cèl·lules. Com hem explicat prèviament, darrere de l’establiment d’un pla de formació del cervell i de la generació de derivats específics hi ha els gens. Per tant, si aconseguim determinar quins gens hi ha actius en cada territori durant el desenvolupament, podríem caracteritzar els límits entre ells i a més definir quants components diferents s’originen a partir de cada regió amb gran precisió. Els estudis de la genoarquitectura neural fan referència a les descripcions de l’estructura neural en termes de patrons d’expressió gènica definits. Ja sabem que la identitat d’un territori la donen les xarxes gèniques que s’estan expressant en una posició i moment determinats durant el desenvolupament. Per tant, l’estratègia és localitzar en la zona d’interès l’ARN missatger (ARNm), que és el producte de la transcripció d’un gen actiu, mitjançant la tècnica d’hibridació in situ (figura 3). Així, la genoarquitectura neural fa referència a la descripció de les estructures neurals en termes de patrons d’expressió gènica, i implica l’ús de les sondes d’ARNm com a marcadors morfològics. Aquests estudis genoarquitectònics permeten una discriminació morfològica que està revolucionant la neuroanatomia moderna (Puelles i Ferran, 2012; Ferran et al., 2015).
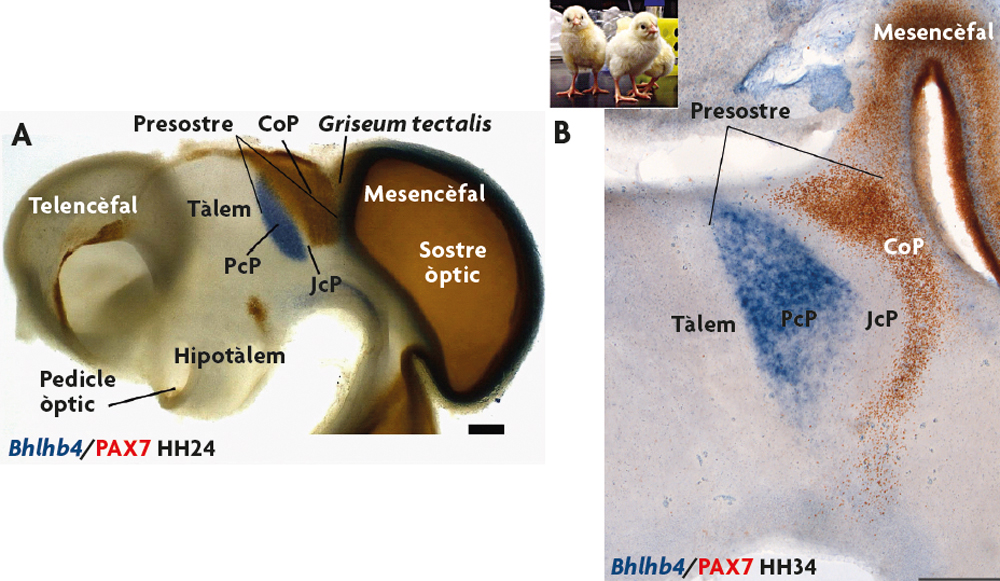
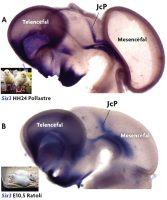
Figura 3. Genoarquitectura cerebral d’aus. A) Vista lateral del cervell d’un pollastre de quatre dies de desenvolupament (estadi embrionari HH24). En l’exemple s’assenyala la regió analitzada (pretectum o presostre), i s’observen dos patrons d’expressió que permeten definir distintes subdivisions o dominis d’expressió (ARNm del gen Bhlhb4, detectat mitjançant hibridació in situ; i proteïna PAX7, detectada mitjançant immunohistoquímica). B) Secció parasagital d’un embrió de pollastre de vuit dies de desenvolupament (HH34). En analitzar els mateixos patrons d’expressió gènica (missatger i proteïna) s’observa que ambdós gens s’expressen en les mateixes subdivisions del cervell que en estadis previs. Mitjançant l’exploració genoarquitectònica es fan evidents tres dominis anteroposteriors de la regió pretectal en tots els estadis analitzats: precomissural (PcP), juxtacomissural (JcP) i comissural (CoP). / José Luis Ferran
Ara bé, els gens s’expressen durant distintes etapes del desenvolupament i de la vida postnatal d’un individu, i a més ho fan en regions anatòmiques diferents. Per tant, únicament la presència del producte d’un gen no és prou per a definir si el territori identificat és comparable o no entre dos estadis de desenvolupament de la mateixa espècie (o entre dues espècies diferents). A més cal conèixer la posició que ocupen aquests productes gènics dins del pla general d’organització del sistema nerviós central (la seua posició topològica). La genoarquitectura és una potent eina que permet caracteritzar des d’estadis molt primerencs del desenvolupament les regions del sistema nerviós, així com les cèl·lules i estructures que en deriven en estadis més tardans i en la vida postnatal. A manera d’exemple, en la figura 3 es mostra una imatge del sistema nerviós central d’un embrió de pollastre, en la qual es pot reconèixer una regió del cervell coneguda com pretectum o presostre que es localitza a la part caudal del diencèfal. Com es pot observar en estats primerencs del desenvolupament, quan aquesta regió té encara poques cèl·lules, es poden identificar diverses subdivisions pels patrons d’expressió de diferents gens (la seua genoarquitectura). Si avancem en el desenvolupament, les cèl·lules que han derivat d’aquestes regions semblen conservar l’expressió d’aquests gens (figures 3 i 4). És a dir, aquesta eina permet distingir cèl·lules amb identitats comunes, i ens proporciona una caracterització anatòmica del cervell molt més elaborada que la que posseíem (figura 4). Mitjançant la genoarquitectura es pot observar amb el màxim detall com són les característiques anatòmiques d’aquesta zona durant tot el desenvolupament embrionari i la vida postnatal d’una espècie (i.e. durant la seua ontogènia) (Ferran, Sánchez-Arrones, Sandoval i Puelles, 2007; Ferran et al., 2009).
«A l’inici del desenvolupament les semblances entre cervells són significatives, ja que partim d’un pla comú, però quan el desenvolupament avança hi ha variacions en les grandàries de les regions o en les característiques de les seues cèl·lules»
D’altra banda, gràcies a l’estudi comparat de l’expressió d’aquests gens durant el desenvolupament es poden identificar regions equivalents en el cervell de distintes espècies de vertebrats (això es coneix com a homologia de camp i les regions comparades es consideren homòlogues; Puelles i Medina, 2002). Però és important insistir que només són comparables les regions de diferents espècies que tenen la mateixa localització dins del pla general (mateixa posició topològica), o els grups de cèl·lules que s’han originat a partir del mateix lloc. En el nostre cas, en la regió pretectal que es mostra en la figura 5, s’observa el mateix gen actiu en la mateixa subdivisió d’aquesta regió tant en pollastre com en ratolí. Això ens revela que aus i mamífers estan usant els mateixos gens en la construcció d’aquesta regió i permet reconèixer la mateixa zona en dues espècies distintes i molt llunyanes evolutivament entre si (Ferran et al., 2008; Merchán, Bardet, Puelles i Ferran, 2011; Morona, Ferran, Puelles i González, 2011).
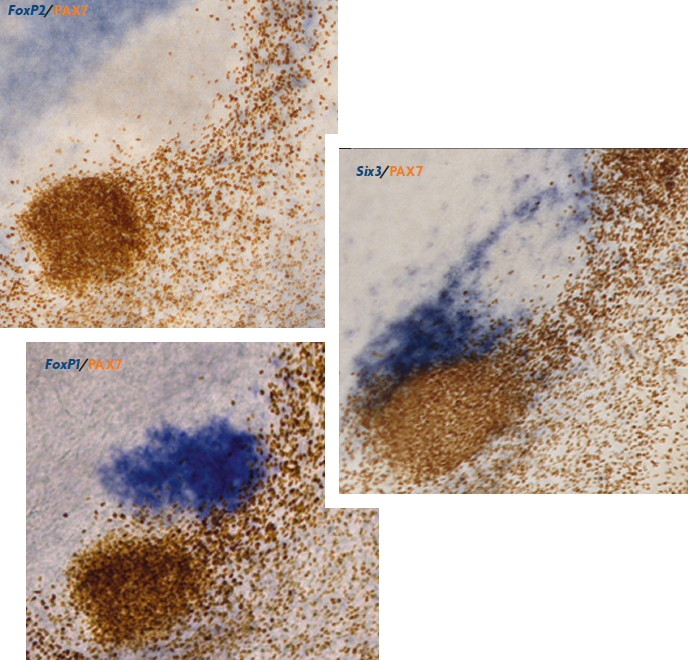
La tasca més difícil és reconèixer homologies quan comparem adults o estadis avançats del desenvolupament, ja que les mateixes regions poden arribar a ser molt diferents en animals adults de distintes espècies, amb gens actius també distints. La dificultat és fins i tot major si considerem que a vegades les cèl·lules migren llargues distàncies a través de diferents dominis, i es fa llavors molt difícil saber el seu lloc d’origen si només s’estudia l’estructura en edat madura. En aquest cas, l’estudi ha de recolzar-se en altres eines que ens indiquen d’on ha derivat l’estructura anatòmica o les cèl·lules d’interès. La raó és que per a considerar dos derivats com a homòlegs han d’haver-se originat a partir del mateix compartiment. Actualment la genoaquitectura es fa servir per a estudiar totes les regions del cervell en distintes espècies, i proporciona detalls sobre l’origen durant el desenvolupament i l’evolució dels distints grups de neurones del cervell inimaginables dècades arrere.
Podem esperar més nivells de complexitat anatòmica?
Hem analitzat que el producte d’un gen, és a dir l’ARN missatger, és una eina summament útil en la caracterització anatòmica de regions del cervell i els seus derivats. Per a tal fi hi ha prou amb l’ús de tècniques que identifiquen aquests productes. No obstant això, avui dia sabem que els ARN missatgers produïts per un únic gen poden ser acoblats de forma diferent, un procés conegut com splicing alternatiu.
Segons aquest mecanisme, un gen pot donar lloc a productes d’ARNm amb diferències quant a la seua seqüència final; això significa que un mateix gen pot produir distintes proteïnes. En relació al nostre estudi, implicaria que dues regions anatòmiques que estan expressant el mateix gen podrien estar produint molècules que no serien exactament iguals i que podrien tenir funcions diferents. La caracterització en alguns casos d’ARNm de splicing alternatiu ha demostrat nivells addicionals en la complexitat de les estructures anatòmiques.
Gens arquitectes del cervell humà
Els productes gènics tenen un paper fonamental en el desenvolupament i funcionament del cervell, però a més poden ajudar-nos a reconèixer quin és el nivell de detall en què el cervell d’un humà s’assembla al d’un altre vertebrat. A l’inici del desenvolupament les semblances entre cervells són significatives, ja que partim d’un pla comú, però quan el desenvolupament avança hi ha variacions en les grandàries de les regions o en les característiques de les seues cèl·lules, i en molts casos apareixen «novetats» evolutives. Conèixer els detalls sobre la formació del cervell d’altres vertebrats ens permetrà avançar en la comprensió de la manera com es forma el cervell humà durant el desenvolupament i quin és el seu origen evolutiu, com també entendre els possibles canvis que podria afrontar el cervell de la nostra espècie en aquest planeta o en altres que poguérem colonitzar en un futur pròxim.
REFERÈNCIES
Davidson, E. H. (2006). The regulatory genome. Burlington, MA: Academic Press.
Ferran, J. L., Sánchez-Arrones, L., Sandoval, J. E., & Puelles, L. (2007). A model of early molecular regionalization in the chicken embryonic pretectum. Journal of Comparative Neurology, 505(4), 379–403. doi: 10.1002/ cne.21493
Ferran, J. L., Sánchez-Arrones, L., Bardet, S. M., Sandoval, J. E., Martínez-de-la-Torre, M., & Puelles, L. (2008). Early pretectal gene expression pattern shows a conserved anteroposterior tripartition in mouse and chicken. Brain Research Bulletin, 75(2-4), 295–298.
Ferran, J. L., de Oliveira, E. D., Merchan, P., Sandoval, J. E., Sánchez-Arrones, L., Martínez-de-la-Torre, M., & Puelles, L., (2009). Genoarchitectonic profile of developing nuclear groups in the chicken pretectum. Journal of Comparative Neurology, 517(4), 405–451. doi: 10.1002/cne.22115
Ferran, J. L., Ayad, A., Merchán, P., Morales-Delgado, N., Sánchez-Arrones, L., Alonso, A., … Puelles, J. (2015). Exploring brain genoarchitecture by single and double chromogenic in situ hybridization (ISH) and immunohistochemistry (IHC) in whole-mount embryos. En G. Hauptmann (Ed.), In situ hybridization methods (pp. 61–82). Nova York: Springer. doi: 10.1007/978-1-4939-2303-8_4
Merchán, P., Bardet, S. M., Puelles, L., & Ferran, J. L. (2011). Comparison of pretectal genoarchitectonic pattern between quail and chicken embryos. Frontiers in Neuroanatomy, 5, 23. doi: 10.3389/fnana.2011.00023
Morona, R., Ferran, J. L., Puelles, L., & González, A. (2011). Embryonic genoarchitecture of the pretectum in Xenopus laevis: A conserved pattern in tetrapods. The Journal of Comparative Neurology, 519(6), 1024–1050. doi: 10.1002/cne.22548
Nieuwenhuys, R., Voogd, J., & Van Huijzen, C. (Eds.). (2008). The human central nervous system. Berlín: Springer. doi: 10.1007/978-3- 540-34686-9
Ohno, S. (1970). Evolution by gene duplication. Londres: Allen and Unwin.
Puelles, L., & Medina, L. (2002). Field homology as way to reconcile genetic and developmental variability with adult homology. Brain Research Bulletin, 57(3–4), 243–255.
Puelles, L., & Ferran, J. L. (2012). Concept of neural genoarchitecture and its genomic fundament. Frontiers in Neuroanatomy, 6, 47. doi: 10.3389/ fnana.2012.00047
Puelles, L., & Rubenstein, J. L. (2003). Forebrain gene expression domains and the evolving prosomeric model. Trends in Neuroscience, 26(9), 469– 476. doi: 10.1016/S0166-2236(03)00234-0
Puelles, L. & Rubenstein J. L., (2015). A new scenario of hypothalamic organization: Rationale of new hypotheses introduced in the updated prosomeric model. Frontiers in Neuroanatomy, 9, 27. doi: 10.3389/fnana.2012.00047
Putnam, N. H., Butts, T., Ferrier, D. E. K., Furlong, R. F., Hellsten, U., Kawashima, T., … Rokshar, D. S. (2008). The amphioxus genome and the evolution of the chordate karyotype. Nature, 453(7198), 1064–1071. doi: 10.1038/ nature06967




