

El cenancestre es defineix com l’últim ancestre comú de tots els éssers vius que existeixen en l’actualitat. La seua naturalesa s’ha inferit a partir de la identificació dels gens homòlegs entre els llinatges d’arqueus, bacteris i eucariotes. Aquestes inferències indiquen que el cenancestre posseïa un sistema de traducció de proteïnes relativament modern i que era semblant en complexitat a una cèl·lula actual. No obstant això, els enzims clau que s’encarreguen tant de la replicació del material genètic com de la biosíntesi de les membranes cel·lulars no són homòlegs entre els bacteris, els arqueus i els eucariotes. Aquí revisem breument la història del concepte de l’últim ancestre comú i les distintes hipòtesis que s’han proposat sobre la seua biologia.Paraules clau: filogènia universal, LUCA, transferència horitzontal, evolució primerenca, ancestre comú.
Els biòlegs evolutius ensopeguen amb el mateix dilema lògic que els historiadors i només poden presentar arguments basats en la suposició que, de totes les seqüències històriques plausibles, n’hi ha una que és més probable que siga una descripció correcta dels fets del passat.
Lynn Margulis, 1975, 29:21-38
El concepte d’homologia i l’arbre universal de la vida
Diuen que una imatge val més que mil paraules. En el cas de la teoria de l’evolució, una de les imatges més suggestives que hi ha es troba en un llibre de notes escrit per Darwin temps abans de la publicació de l’Origen de les espècies (figura 1). En aquest quadern es pot apreciar que Darwin representa mitjançant un dibuix l’origen de distintes espècies a partir d’un ancestre comú.
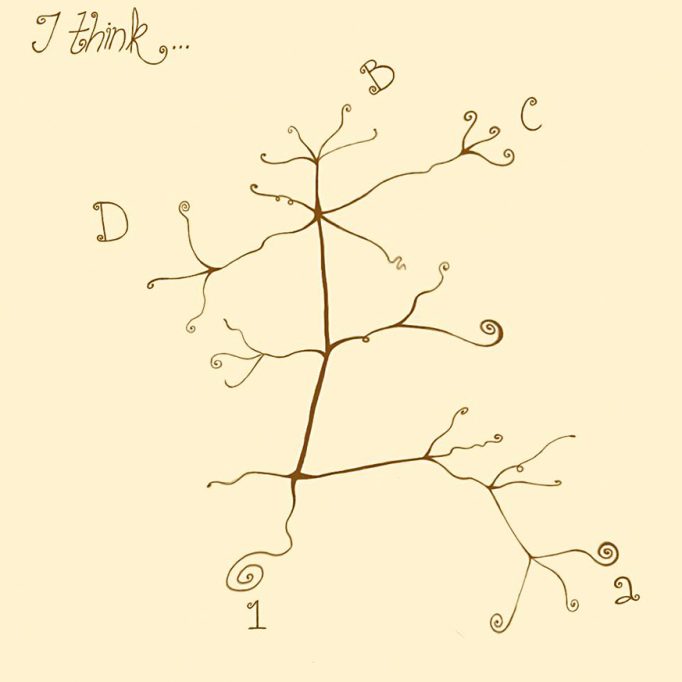
Figura 1. Darwin va esquematitzar l’evolució d’un grup d’espècies a partir d’una espècie ancestral en el seu quadern de notes conegut com «Notebook B». / Dibuix de Sofía Delaye Pascual
Com veiem, la teoria de l’evolució de Darwin suggereix que les distintes espècies que hi ha avui dia han evolucionat (diversificat) les unes de les altres a partir d’ancestres comuns. Aquesta idea, tan simple en principi, va canviar profundament la manera d’entendre els éssers vius. Per exemple, quan comparem les extremitats de distints mamífers, trobem que els ossos que les conformen són semblants entre si (figura 2). D’acord amb Darwin, aquesta similitud s’explica perquè aquestes extremitats van evolucionar a partir d’un ancestre comú, és a dir, són homòlogues. De fet, Darwin va escriure en L’origen (1859, p. 415):
Què podria ser més curiós que el fet que la mà de l’home, formada per a agafar coses; la del talp, per a cavar; la pota del cavall, l’aleta del marsuí i l’ala de la ratapinyada estiguin totes construïdes segons el mateix patró […]?
El concepte d’homologia ens permet inferir part de les característiques que posseïa l’ancestre comú d’un grup determinat d’organismes. Per exemple, en el cas dels mamífers de la figura 2 podem inferir que l’avantpassat comú a tots ells tenia aquests ossos en les mateixes posicions relatives. La teoria de Darwin obre la possibilitat que tots els éssers vius estiguen relacionats en un gran arbre universal de la vida per haver evolucionat a partir d’un únic ancestre comú. De fet, Darwin va suggerir en L’origen (1859, p. 455): «[…] probablement, tots els éssers orgànics que mai han viscut en aquest món han descendit d’alguna forma primordial, en la qual la vida fou insuflada per primera vegada.»
No obstant això, Darwin, previngut com era en les seues asseveracions científiques, no va gosar mai de representar l’evolució de tots els éssers vius en un sol arbre. Va ser Ernst Haeckel qui va realitzar un dels primers dibuixos d’un arbre universal. Com podem apreciar en la figura 3, Haeckel suggereix que els éssers vius divergim en plantes, animals i protists a partir d’un tronc comú.
«La teoria de Darwin obre la possibilitat que tots els éssers vius estiguem relacionats en un gran arbre universal de la vida per haver evolucionat a partir d’un únic ancestre comú»
Posteriorment, durant la primera meitat del segle xx, Chatton (1938) i amb més claredat Stanier i Van Niel (1941) van proposar que els éssers vius s’han de classificar en dos grups principals: procariotes i eucariotes (figura 4). Els eucariotes es defineixen per posseir membranes internes, com ara la membrana nuclear i el reticle endoplàsmic, i per dividir-se per mitosi, i els procariotes per no tenir membranes internes i dividir-se per fissió binària. En paraules de Stanier i Van Niel (1941, p. 464): «[…] el regne Monera, compost de microorganismes sense un vertader nucli ni plastidis ni reproducció sexual.»
El 1969, l’ecòleg Robert Whittaker proposa que els éssers vius s’han de classificar en quatre grans regnes eucariotes (Plantae, Animalia, Protista i Fungi) i un de procariota (Monera). Aquest esquema de classificació va ser adaptat per Lynn Margulis i combinat amb la divisió procariota/eucariota per a acomodar l’origen de les mitocondris i els cloroplasts a partir dels alfa proteobacteris i cianobacteris respectivament (figura 5). No obstant això, va ser amb l’adveniment de la biologia molecular quan la naturalesa de l’últim ancestre comú va poder començar a ser tema directe d’investigació.
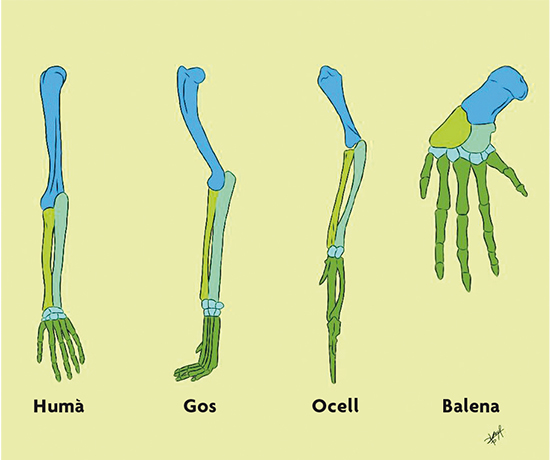
Figura 2. Extremitats homòlogues entre distints mamífers. / Dibuix de Sofía Delaye Pascual
L’ancestre universal, la història del concepte
A mitjan segle xx Frederick Sanger va desenvolupar les tècniques necessàries per a conèixer les seqüències d’aminoàcids i nucleòtids que conformen les proteïnes i el DNA respectivament. La informació generada per aquestes tècniques va permetre a Emile Zuckerkandl i Linus Pauling suggerir que era possible reconstruir la història dels éssers vius comparant aquestes biomolècules.
El 1977, utilitzant tècniques moleculars, es va publicar un dels descobriments més sorprenents sobre la diversitat de la vida a la Terra. Carl Woese i George Fox van descobrir que, d’acord amb les similituds i diferències en la subunitat petita de la molècula de l’RNA ribosòmic (SSU rRNA en les sigles en anglès), els éssers vius ens dividim en tres grups (Woese i Fox, 1977a). Aquests tres grups (o llinatges) són: (a) els eubacteris, (b) el nucli-citoplasma eucariota, i (c) un grup de procariotes que van denominar com arqueobacteris. En l’actualitat aquests tres grups es coneixen com a Bacteris, Eucariotes i Arqueus.
Basant-se en aquesta divisió universal, Woese i Fox van suggerir que en la divergència dels tres llinatges cel·lulars existia una entitat primitiva en què la relació entre genotip i fenotip encara no havia evolucionat a la seua forma actual (Woese i Fox, 1977b, p. 1). Aquesta entitat biològica ancestral la denominaren progenot: «Aquesta entitat primitiva s’ha denominat progenot, terme que mostra la possibilitat que encara no s’haja completat l’evolució de la relació entre genotip i fenotip.» Alguns anys més avant, Fitch i Upper (1987, p. 761), en estudiar l’evolució del codi genètic, encunyen el terme cenancestre i el defineixen com: «L’antecessor comú més recent de tots els organismes que són vius avui.»
«Va ser amb l’adveniment de la biologia molecular quan la naturalesa de l’últim ancestre comú va poder començar a ser tema directe d’investigació»
És important notar les diferències entre ambdós conceptes. El progenot implica un estat primitiu, en tant que el concepte de cenancestre no necessàriament. El cenancestre, també conegut com «Last Universal Common Ancestor» o LUCA, és només l’últim ancestre comú de tots els éssers vius que existeixen actualment i aquest ben bé podia haver estat simple com un progenot o complex com una cèl·lula actual.
Gràcies al desenvolupament de la tecnologia de seqüenciació del DNA i a l’acumulació d’una gran quantitat i diversitat de seqüències en les bases de dades, va ser possible començar a identificar els gens conservats entre els Bacteris, Arqueus i Eucariotes (figura 6). Això va permetre aplicar una metodologia simple per a inferir els gens que en principi es trobaven presents en el genoma del cenancestre. Basant-se en aquesta metodologia, Lazcano, Fox i Oró proposen (1992) que l’últim ancestre comú posseïa una maquinària genètica semblant a la d’una cèl·lula procariota actual.
No obstant això, la filogènia proposada per Woese i Fox no té arrel. És a dir, no mostra si algun dels llinatges cel·lulars és més antic que els altres dos. La primera aproximació per a tractar d’identificar la posició de l’arrel de l’arbre universal va ser realitzada independentment per dos grups d’investigació en utilitzar gens duplicats universalment conservats. Ambdues anàlisis suggerien que l’arrel de l’arbre universal es troba en la branca dels bacteris (figura 7). Això va reforçar la idea que el cenancestre era de naturalesa procariota.
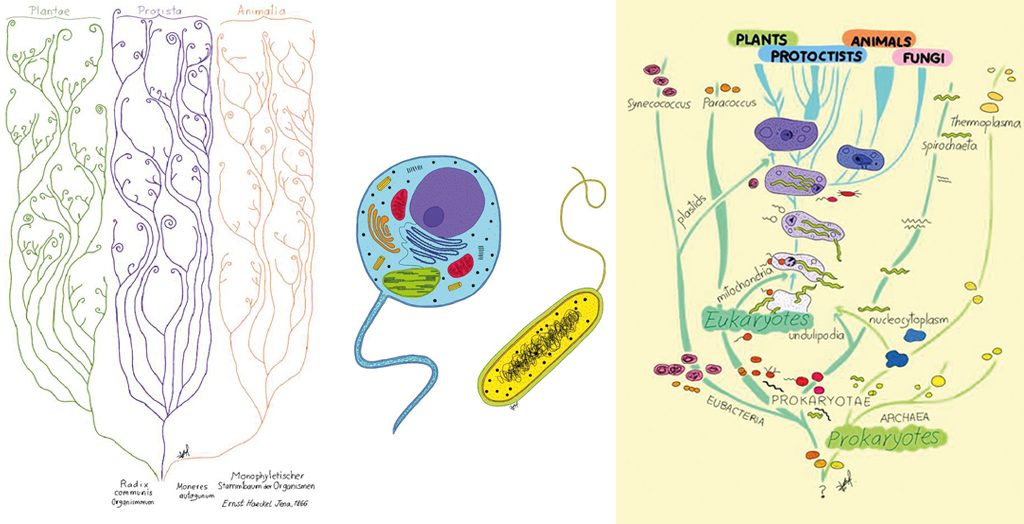
A l’esquerra, figura 3, arbre de la vida inspirat en Generelle Morphologie der Organismen (1866), d’Ernst Haeckel. Al mig, figura 4, observem que a escala cel·lular podem classificar tots els éssers vius en procariotes i en eucariotes. A la dreta, figura 5, esquema de classificació dels cinc regnes modificat per Lynn Margulis (1996). En la imatge observem la divisió citològica dels éssers vius en procariotes i eucariotes. També mostra que els procariotes (Arqueus i Bacteris) donen origen als eucariotes (Animals, Plantes, Fongs i Protistes) per simbiogènesi. / Dibuix de Sofía Delaye Pascual
RNA o DNA?
El 1996 Mushegian i Koonin van comparar els genomes dels bacteris Haemophilus influenzae i Mycoplasma genitalium per proposar el conjunt mínim i suficient de gens necessaris per a sustentar la vida cel·lular. També van buscar gens homòlegs a H. influenzae i M. genitalium tant en Arqueus com en Eucariotes per tractar d’identificar gens universalment conservats i inferir la naturalesa de l’últim avantpassat comú. Com que no van trobar gens homòlegs per a l’enzim que s’encarrega de replicar el DNA en els tres llinatges cel·lulars, Mushegian i Koonin van proposar que l’últim ancestre comú dels éssers vius posseïa un genoma d’RNA. Això significaria que l’últim ancestre comú seria més simple que qualsevol cèl·lula coneguda avui dia. No obstant això, inferències posteriors han mostrat que el conjunt de gens conservats no és consistent amb el nivell de complexitat d’un progenot.
La transferència horitzontal de gens i l’arbre de la vida
Una de les principals sorpreses que va comportar la seqüenciació de genomes complets a finals de la dècada dels noranta va ser la freqüent falta de concordança de la filogènia proposada per l’rRNA, en què els éssers vius s’agrupen en Arqueus, Bacteris i Eucariotes, amb les filogènies derivades d’altres gens conservats en aquests tres llinatges cel·lulars. Si bé a l’inici es va pensar que moltes de les discrepàncies es devien a problemes metodològics, estudis posteriors van demostrar que, almenys entre procariotes, una proporció important de les incongruències filogenètiques es devien al fenomen de la transferència horitzontal de gens (figura 8).
El descobriment de l’abundància d’esdeveniments de transferència horitzontal al llarg de la història de la vida en la Terra va plantejar un problema important per a la reconstrucció del conjunt de gens presents en l’últim ancestre comú. De fet, s’ha arribat a proposar que durant les etapes primerenques de l’evolució de la vida en la Terra l’herència horitzontal era el mode més comú d’herència (Woese, 1998).
Gens nous, gens perduts i el cenancestre
D’una banda, un genoma pot guanyar gens ja siga per transferència horitzontal o per creació de novo. D’altra banda, un gen que ja no contribueix a la supervivència d’un organisme pot perdre’s al llarg de l’evolució. Com ja vam veure, la freqüència d’aquests esdeveniments afecta la reconstrucció del contingut gènic de l’últim ancestre comú. En un intent per tractar d’inferir el contingut de gens del cenancestre tenint en compte els esdeveniments de guany i pèrdua de gens al llarg de la història de la vida en la Terra, Mirkin i col·laboradors (2003) van desenvolupar un algoritme basat en l’argument de la parsimònia. D’acord amb el seu algoritme, l’últim ancestre comú tindria al voltant de 572 gens. Aquest conjunt de gens és quasi suficient per a codificar un metabolisme coherent. No obstant això, hi ha dues absències importants. La primera, com ja mencionem anteriorment, és la falta d’una polimerasa de DNA. La segona gran absència són dos enzims clau encarregats de la síntesi de lípids de la membrana cel·lular.
La falta d’aquests components és sorprenent si considerem que representen dues de les tres propietats més importants dels éssers vius: la delimitació d’un ésser viu del seu ambient a través d’una membrana plasmàtica i l’herència del material genètic basada en la replicació d’un àcid nucleic (Szathmáry, 2005). L’altra característica important dels éssers vius, de la qual parla Eörs Szathmáry i que sí que és present en la reconstrucció de Mirkin, és el metabolisme. La disputa sobre la naturalesa del genoma de l’últim avantpassat comú i la naturalesa de les seues membranes encara no està tancada en la comunitat científica (Peretó, López-García i Moreira, 2004; Poole et al., 2014).
L’últim ancestre comú i els virus
Un altre aspecte no clar sobre la naturalesa del cenancestre és si els virus existien en la seua època i si aquest era susceptible a ser «infectat». Fins al dia d’avui no comptem amb evidència decisiva que manifeste la coexistència de virus amb cèl·lules tan antigues com l’ancestre comú. No obstant això, diferents hipòtesis proposen que aquestes entitats no sols van coincidir, sinó que els virus van precedir les cèl·lules, o fins i tot que van tenir un paper central en etapes molt primerenques de la vida, pròximes al seu origen (Agol, 2010).
Hi ha moltes preguntes sense resoldre sobre l’origen i l’evolució dels virus. No obstant això, molts investigadors considerem que aquestes entitats biològiques són d’origen polifilètic. Per tant, es fa molt difícil o impossible provar-ne la presència en etapes primerenques de la vida, a causa de la seua naturalesa de material «escapat» d’organismes cel·lulars. Aquesta posició, però, és lluny de suscitar un consens ampli i planteja una sèrie d’objeccions. Per exemple, el fet que unes poques, però importants, estructures proteiques com el jelly-roll de la càpside estiguen presents en diversos tipus virals, suggereix un origen comú. Addicionalment, s’ha proposat una llista breu de gens possiblement homòlegs a diversos grups de virus, els anomenats hallmark genes (Koonin i Dolja, 2013), abonant la proposta monofilètica. Si fos cert, aquestes dades serien evidència d’un origen comú per a tots els virus, i suggeriria que el dit ancestre viral (o ancestres) va ser contemporani o almenys pròxim a l’ancestre cel·lular.
És important recalcar que la informació obtinguda fins al moment no permet discutir la presència dels virus en èpoques pròximes al cenacestre i no es poden extrapolar les dades obtingudes com a evidència d’etapes més primerenques de la vida o encara menys proposar que els virus van tenir un paper central en el pas d’organismes amb genoma d’RNA a DNA o fins i tot en l’origen de la vida. Els mètodes moleculars i la comparació de seqüències no poden extraure informació d’etapes precel·lulars.
Tot i això, si l’últim ancestre comú era ja un organisme tan complex com una cèl·lula procariota i si els virus són essencialment material «escapat» d’una maquinària genètica complexa, tampoc podem descartar la presència de virus en una època pròxima al cenancestre. Saber si el nostre ancestre universal patia infeccions virals o almenys coevolucionava amb els virus és un camp que requereix de més estudis igual com moltes de les preguntes que ens presenta l’estudi de l’evolució primerenca de la vida.
Conclusions
Hi ha un fil que ens connecta des dels nostres dies fins als primers éssers vius. Aquest fil és format per incomptables generacions d’ancestres i descendents modificats. Generacions que en conjunt donen forma a l’arbre universal de la vida en la Terra que alguna vegada va imaginar Darwin. Les nostres inferències sobre la biologia del cenancestre estan íntimament lligades a l’estructura d’aquest arbre universal i als processos evolutius que li donen forma.
REFERÈNCIES
Agol, V. I. (2010). Which came first, the virus or the cell? Paleontological Journal, 44(7), 728–736. doi: 10.1134/S0031030110070038
Chatton, E. (1938). Titre et travaux scientifiques (1906–1937) de EdouardChatton. Sète: Sottano.
Darwin, Ch. (1859). On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. Londres: John Murray.
Fitch, W. M., & Upper, K. (1987). The phylogeny of tRNA sequences provides evidence of ambiguity reduction in the origin of the genetic code. Cold Spring Harbor Symposia on Quantitative Biology, 52, 759–767. doi: 10.1101/SQB.1987.052.01.085
Koonin, E. V., & Dolja, V. V. (2013). A virocentric perspective on the evolution of life. Current Opinion in Virology, 3, 546–557. doi: 10.1016/j.coviro.2013.06.008
Lazcano, A., Fox, G. E., & Oró, J. (1992). Life before DNA: The origin and evolution of early Archean cells. En R. P. Mortlock (Ed.). The evolution of metabolic function (pp. 237–295). Boca Raton: CRC Press.
Margulis, L. (1975). Symbiotic theory of the origin of eukaryotic organelles: Criteria for proof. Symposia of the Society for Experimental Biology, 29, 21–38.
Mirkin, B. G., Fenner, T. I., Galperin, M. Y., & Koonin, E. V. (2003). Algorithms for computing parsimonious evolutionary scenarios for genome evolution, the last universal common ancestor and dominance of horizontal gene transfer in the evolution of prokaryotes. BMC Evolutionary Biology, 3(2). doi: 10.1186/1471-2148-3-2
Peretó, J., López-García, P., & Moreira, D. (2004). Ancestral lipid biosynthesis and early membrane evolution. Trends in Biochemical Sciences, 29(9), 469–477. doi: 10.1016/j.tibs.2004.07.002
Poole, A. M., Horinouchi, N., Catchpole, R. J., Si, D., Hibi, M., Tanaka, K., & Ogawa, J. (2014). The case for an early biological origin of DNA. Journal of Molecular Evolution, 79 (5–6), 204–212. doi: 10.1007/s00239-014-9656-6
Stanier, R. Y., & Van Niel, C. B. (1941). The main outlines of bacterial classification. Journal of Bacteriology, 42(4), 437–463.
Szathmáry, E. (2005). Life: In search of the simplest cell. Nature, 433, 469-470. doi: 10.1038/433469a
Whittaker, R. H. (1969). New concepts of kingdoms of organisms. Science, 163, 150–160. doi: 10.1126/science.163.3863.150
Woese, C. R., & Fox, G. E. (1977a). Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proceedings of the National Academy of Sciences, 74, 5088–5090. doi: 10.1073/pnas.74.11.5088
Woese, C. R., & Fox, G. E. (1977b). The concept of cellular evolution. Journal of Molecular Evolution, 10, 1–6. doi: 0.1007/BF01796132
Woese, C. R. (1998). The universal ancestor. Proceedings of the National Academy of Sciences, 95, 6854–6859.




