

L’evolució del gènere ‘Homo’
Noves incògnites i perspectives

En aquest treball es passa revista a les principals qüestions que afecten l’evolució del gènere Homo, com és el cas del seu origen, el problema de la variabilitat en Homo erectus i l’impacte de la paleogenòmica. Encara no s’ha arribat a un consens sobre el candidat australopitecí que va donar lloc als primers representants assignables a Homo. Aquesta discussió afecta fins i tot el reconeixement mateix de les espècies H. habilis i H. rudolfensis. Pel que fa a la qüestió de la variabilitat dels primers paleodemes assignats a Homo, les troballes fetes al jaciment georgià de Dmanisi han posat en dubte alguns dels criteris utilitzats fins ara per distingir entre espècies com ara H. erectus o H. ergaster. Finalment, la irrupció de la paleogenòmica ha posat en evidència que el flux genètic entre antigues poblacions d’hominins va ser major que no s’esperava.
Paraules clau: paleogenòmica, gènere Homo, hominins, variabilitat, Dmanisi.
En els últims anys, la nostra concepció de l’origen i evolució del nostre gènere s’ha vist sacsejada per diferents troballes que, lluny de donar resposta als problemes que es formulaven a finals del segle XX, han reobert debats i han obligat a replantejar-se els esquemes que es van mantenir vigents durant dècades. Algunes d’aquestes qüestions obertes ho són perquè falten els fòssils adequats que ens donen la resposta. D’altres, per contra, són encara objecte de debat per la diferent interpretació que es dóna a alguns fòssils i a l’adscripció sistemàtica d’aquests. Troballes com les del jaciment georgià de Dmanisi o les proporcionades per la paleogenòmica posen en dubte, sobretot, els criteris utilitzats fins ara per al reconeixement d’espècies en el registre fòssil. Tot seguit enumerem algunes d’aquestes qüestions.
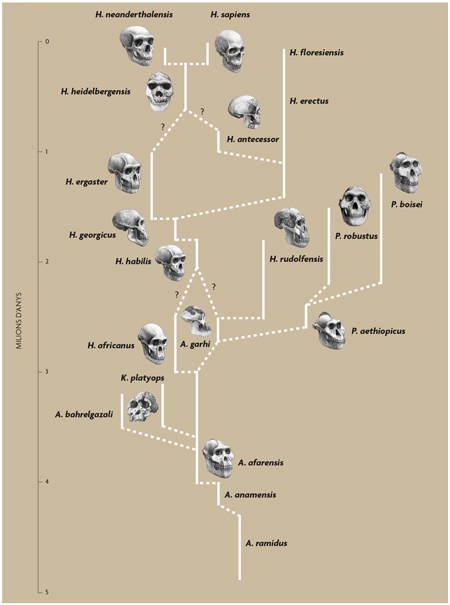
Esquema de l’evolució humana proposat per Agustí i Lordkipanidze (2005). En els últims anys, la nostra concepció sobre l’origen i evolució del gènere Homo s’ha vist sacsejada per diferents troballes que han reobert debats i obligat a replantejar-se determinats esquemes. / Il·lustracions dels cranis: Mauricio Antón
L’origen del gènere ‘Homo’
Els primers representants del nostre gènere fan aparició en el registre fòssil africà fa uns 2,8 milions d’anys (Villmoare et al., 2015). Aquestes primeres restes del nostre gènere han estat atribuïdes de vegades a Homo rudolfensis. Tenim evidències d’una segona espècie, Homo habilis de fa un poc més de dos milions d’anys, que es diferencia de l’anterior en alguns trets secundaris i en la menor capacitat craniana, encara que per a alguns investigadors Homo habilis i Homo rudolfensis corresponen en realitat a la mateixa espècie.
Fins a mitjan anys setanta del passat segle hi havia un clar candidat australopitecí per a ocupar el lloc d’avantpassat del nostre gènere, Australopithecus africanus, de distribució exclusivament sud-africana. La publicació el 1978 de les restes d’Australopithecus afarensis procedents d’Afar, a Etiòpia, va representar el primer qüestionament d’aquesta filiació. Per als seus descobridors, Don Johanson i Tim White, Australopithecus afarensis, amb una cronologia entre 3,5 i 3 milions d’anys, encaixava com a perfecte antecessor del nostre gènere, mentre que Australopithecus africanus probablement va donar lloc a Sud-àfrica a les formes robustes d’australopitecins enquadrades en el gènere Paranthropus.
Amb posterioritat, el 1996, el mateix White va proposar una nova espècie, Australopithecus garhi, trobada en nivells datats en uns 2,5 milions d’anys a la localitat de Bouri, a Etiòpia. Segons White i els seus col·laboradors, A. garhi es trobava en el lloc i en el moment adequat per a ser l’avantpassat del nostre gènere. S’hi va afegir el fet d’haver trobat associats a A. garhi ossos que presentaven marques de tall d’estris lítics, dels quals se suposa que aquesta espècie va ser la responsable.
Per a acabar de complicar el tema, el 2001 l’equip liderat per Meave Leakey va proposar un nou gènere, Kenyanthropus, per a un crani molt deformat procedent de la conca de Lomekwi a Kenya, i datat en uns 3,5 milions d’anys (és a dir, contemporani d’A. afarensis). La capacitat cranial estimada de l’anomenat Kenyanthropus platyops era baixa, entorn de 350 cm3. Però el que va portar els seus descobridors a proposar un nou gènere va ser la cara, llarga i plana. Fent un pas més enllà, van plantejar que aquesta forma era l’avantpassat d’Homo rudolfensis, espècie que van proposar incloure també en el gènere Kenyanthropus. Aquesta última proposició ha trobat escàs eco entre la comunitat paleoantropològica, que, lluny d’acceptar el nou gènere, ha optat per incloure l’espècie K. platyops dins d’Homo o d’Australopithecus. No obstant això, alguns autors sí que han acceptat el suggeriment d’una relació entre K. platyops i H. rudolfensis.
Més enllà de les troballes en l’est d’Àfrica, la idea d’un origen sud-africà del nostre gènere va tornar a eixir a la palestra amb el descobriment d’una nova espècie d’Australopithecus, A. sediba, a la cova de Malapa a Sud-àfrica. Es tracta de dos esquelets parcials pertanyents a una dona jove d’uns vint anys i a un adolescent d’uns dotze. L’element més significatiu és un crani en excel·lent estat de conservació que revela un Australopithecus pròxim a Australopithecus africanus però amb caràcters més derivats. A part que posseïa un cervell una mica més gran que el del seu predecessor, la cara mostra alguns trets que l’aproximen als primers representants del nostre gènere, com ara Homo habilis (per exemple, la forma de la cavitat nasal i els pòmuls). Per al seu descobridor, Lee Berger, de la Universitat de Witwatersrand (Sud-àfrica), Australopithecus sediba constitueix la perfecta anella intermèdia entre Australopithecus africanus i els primers representants del gènere Homo. El problema, però, és que les restes d’Australopithecus sediba estan datades en uns 1,8 milions d’anys i són, per tant, molt posteriors a l’eclosió del nostre gènere, que data de més de 2,5 milions d’anys. A aquest mateix equip és degut el reconeixement d’una nova espècie arcaica del gènere Homo, H. naledi, a Dinaledi Chamber, una cavitat de molt difícil accés també a Sud-àfrica. No obstant això, l’absència d’un context geològic i paleontològic adequat qüestiona la possible rellevància d’aquesta troballa en relació amb l’origen del nostre gènere.

Homo rudolfensis ha estat considerat com el primer representant del gènere Homo, que es remuntaria a fa uns 2,8 milions d’anys. Fa poc més de dos milions d’anys, tindríem evidències de l’existència d’una segona espècie, Homo habilis, encara que alguns investigadors consideren que tant H. rudolfensis com H. habilis són la mateixa espècie. En la imatge, crani i reconstrucció d’Homo rudolfensis (segons Mauricio Antón). / Il·lustració: Mauricio Antón
Pel que fa als candidats australopitecins de l’est d’Àfrica, hi ha una dada sorprenent i és la seua tendència a posseir grans molars (un fenomen conegut com «megadòncia»), que arribarà al màxim exponent amb les espècies incloses en el gènere Paranthropus. Aquesta tendència és ja evident en Australopithecus afarensis, espècie que, si bé podria semblar un bon avantpassat d’Homo, igualment encaixaria com un plausible avantpassat de Paranthropus. De fet, quan va eixir a la llum l’anomenat «crani negre» de Paranthropus aethiopicus, diversos autors van fer notar les seues afinitats amb Australopithecus afarensis. Si anem ara a Australopithecus garhi, aquesta tendència a la megadòncia es fa encara més patent, per bé que la morfologia cranial no permeta incloure-la en Paranthropus. No obstant això, és evident que l’est d’Àfrica és un context ambiental que afavoria el desenvolupament d’un potent aparell mastegador i grans molars.
Quan tot seguit passem a analitzar la dentició dels primers membres del nostre gènere, ens trobem amb la sorprenent conclusió que també les primeres restes assignades a Homo posseïen grans molars, raó per la qual són comunament incloses dins de l’espècie Homo rudolfensis. Només més tardanament apareixen les restes assignades a Homo habilis, la dentició de les quals era més gràcil i pròxima a la dels Australopithecus sud-africans.
Estem, per tant, davant d’una dicotomia. Així, a l’Àfrica oriental podríem reconèixer un llinatge que, a través d’Australopithecus afarensis i Australopithecus ghari (i pot ser Kenyanthropus), condueix a Homo rudolfensis. D’altra banda, hi deu haver una línia gràcil que des d’Australopithecus africanus portaria a Homo habilis, tal vegada incloent-hi Australopithecus sediba. La sorprenent conclusió seria un origen difilètic per al gènere Homo.
Ara bé, davant d’un registre tan fragmentari com el corresponent a les primeres variants del gènere Homo, cal preguntar-se si en realitat tant Homo habilis com Homo rudolfensis corresponen a variants dins d’una mateixa espècie. Els elements diferencials entre ambdós poden ser una conseqüència de la mateixa variabilitat poblacional dins d’aquella espècie.
L’estatus d’‘Homo erectus’ i el tema de la variabilitat
A mitjan anys setanta del segle passat, dues revolucions conceptuals en el camp de la biologia evolutiva influirien profundament en les idees sobre la nostra evolució. D’una part, en la sistemàtica paleontològica es va estendre el recurs a la cladística a l’hora d’establir la filogènia de qualsevol grup fòssil. D’aquesta manera, amb la seua distinció entre caràcters primitius i derivats, es van poder elaborar hipòtesis contrastables sobre el parentiu entre diferents tàxons.

En 2001, l’equip de la paleoantropòloga Meave Leakey va proposar el nou gènere Kenyanthropus. Per a aquest grup d’investigació, Kenyanthropus platyops era l’avantpassat d’Homo rudolfensis, proposta que ha comptat amb escàs eco en la comunitat científica. En la imatge, crani i reconstrucció de Kenyanthropus platyops (segons Mauricio Antón). / Il·lustració: Mauricio Antón
Més influent encara va ser la proposta dels paleontòlegs Niles Eldredge i Stephen J. Gould, segons els quals bona part de les seqüències evolutives no s’adequaven al model simplista de canvi gradual, sinó que les espècies apareixien abruptament durant moments de canvi sobtat i amb posterioritat es mantenien estables fins a l’extinció. En el camp de l’evolució humana, el guant va ser arreplegat per Niles Eldredge i Ian Tattersall, que el 1982 van publicar The myths of human evolution (“Els mites de l’evolució humana”), una obra que tindria una forta influència en la nostra concepció dels patrons evolutius en l’evolució humana. A partir d’ací, el reconeixement que l’evolució humana havia estat un procés molt més variat i complex es va fer sentir en la interpretació dels antics fòssils i d’altres de nous que anaven apareixent. El panorama de l’evolució humana va canviar sobtadament i, sota el nou paradigma, noves espècies van eixir a la palestra: Homo rudolfensis, Homo ergaster, Kenyanthropus platyops, Homo antecessor, Homo georgicus i altres més. Algunes espècies anteriors es van volatilitzar literalment, com el clàssic Homo erectus, que va deixar de ser una anella necessària en l’evolució humana i va quedar restringida a l’entorn asiàtic. La vigència d’aquest paradigma es va mantenir durant els anys vuitanta i part dels noranta del passat segle. A final d’aquesta dècada, no obstant això, s’observen els primers senyals de desacord per part, per exemple, de Tim White, el qual qüestiona la distinció entre Homo erectus i Homo ergaster. Però són sobretot les troballes en el jaciment georgià de Dmanisi les que provocaran novament un canvi en el paradigma imperant.
El jaciment de Dmanisi, datat per mètodes radiomètrics i de paleomagnetisme en 1,8 milions d’anys, ha proporcionat al llarg de les últimes dècades el millor registre fòssil dels primers hominins que van eixir d’Àfrica (Agustí i Lordkipanidze, 2005). Tot va començar el 1991, quan la primera resta humana de Dmanisi, una mandíbula que conservava tota la seua dentició, va eixir a la llum. Des d’un principi aquesta mandíbula va ser objecte de debat, ja que alguns autors van qüestionar la seua antiguitat basant-se en el grau de reducció dels molars posteriors i la situaven en el plistocè mitjà i no en el plistocè inferior. Calgué esperar fins a 1999 perquè la troballa de dos nous cranis confirmara el caràcter arcaic dels hominins de Dmanisi. El primer, D2280, només conservava la calota craniana, és a dir, la volta del crani que allotja el cervell, la qual cosa permetia inferir una capacitat craniana de 775 cm3. El segon, D2282, molt deformat, conservava a més part de la cara i la seua capacitat craniana va poder ser estimada en uns 650 cm3. L’any 2000 va aparèixer una nova mandíbula, aquesta vegada de grans dimensions i amb molars posteriors molt desenvolupats, com corresponia a un representant arcaic del gènere Homo. Altres trets de la mandíbula evidenciaven un clar primitivisme, per la qual cosa el paleontòleg georgià Lee Gabunia i el seu equip van decidir erigir una nova espècie, Homo georgicus. Aquesta mandíbula va donar lloc a les primeres especulacions relatives a la presència de dues espècies d’hominins a Dmanisi.
L’any següent, el 2001, un nou crani va acudir a la cita a Dmanisi. Aquesta vegada, el crani D2700 incloïa la mandíbula i es trobava en un estat quasi perfecte de conservació. L’anàlisi detallada d’aquest crani denotava un hominí molt arcaic, amb una capacitat craniana de 600 cm3. El perfil era còncau i, a diferència del primer crani D2280, presentava arcs superciliars poc desenvolupats. La mandíbula era pareguda a la primera mandíbula de l’any 1991 (Vekua et al., 2002). El crani D2700 va demostrar clarament que els hominins de Dmanisi no podien ser assignats a l’espècie africana Homo ergaster i que la primera eixida d’Àfrica va ser protagonitzada per poblacions més pròximes a Homo habilis que a aquesta última espècie. Aquesta atribució va ser confirmada el 2002, quan va eixir a la llum un nou crani senil, un individu desdentat (la mandíbula corresponent va aparèixer l’any següent, el 2003) que, no obstant això, manifestava els mateixos trets arcaics que el crani del 2001 (Lordkipanidze et al., 2005). La campanya de l’any 2005, finalment, va donar lloc a l’espectacular troballa del crani D4500, que encaixava amb la gran mandíbula de l’any 2000. Aquest crani no va defraudar les expectatives creades per la mandíbula al seu dia. Encara que la seua capacitat craniana era la més baixa de tota la població de Dmanisi, tan sols 546 cm3, mostrava una robustesa que superava la resta de troballes anteriors (Lordkipanidze et al., 2013).
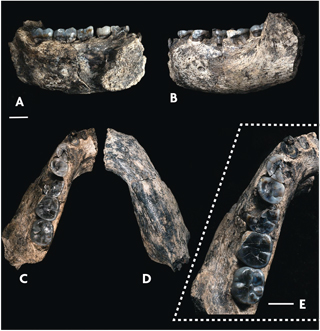
Vista del fòssil d’una mandíbula des de diferents perspectives. / Villmoare et al., 2015
L’anàlisi del paleodeme complet de Dmanisi va portar l’equip paleoantropològic a la conclusió que, a pesar de la seua variabilitat, el conjunt corresponia a una única població. L’anàlisi geològica, paleomagnètica i faunística del jaciment confirmava aquest punt de vista: totes les restes de Dmanisi corresponien a un únic episodi de sedimentació. Aquests arguments van ser contestats per aquells que, en el paleodeme de Dmanisi, hi reconeixien més d’una espècie, i que sense coneixement de causa van qüestionar l’estratigrafia del jaciment. Per als autors de l’estudi del nou crani, però, no hi havia dubte: es tractava d’una població arcaica del nostre gènere que, a pesar de la seua limitada capacitat craniana, mostrava ja els caràcters derivats d’Homo erectus.
La resistència a acceptar que el conjunt de Dmanisi correspon a una única població coetània és comprensible, perquè representa revisar bona part dels criteris utilitzats a l’Àfrica i altres enclavaments per a interpretar el registre fòssil humà. La frontera entre Homo erectus i Homo ergaster, mantinguda durant anys, es dilueix, el mateix que la distinció entre Homo habilis i Homo rudolfensis. El cas és que, a diferència de les múltiples troballes africanes, el paleodeme de Dmanisi correspon a una única població que, amb cinc cranis i restes postcranials de més de tres individus, permet fer-se una idea de la variabilitat real de les primeres poblacions del gènere Homo (Lordkipanidze et al, 2007).
La irrupció de la paleogenòmica
Des de finals del segle XX fins a l’actualitat, la nostra interpretació del registre fòssil s’ha vist sacsejada per l’eclosió d’un nou tipus de fòssil, el DNA fòssil, un tipus de registre inimaginable fa tan sols unes dècades. La seqüenciació de DNA mitocondrial en organismes del plistocè superior va començar amb èxit en la dècada dels vuitanta del segle XX i la seua aplicació als neandertals, amb cronologies que entraven dins del rang de conservació d’aquest material genètic, estava cantada. Curiosament, entre les primeres restes neandertals a ser analitzades, i que van demostrar un acceptable nivell de conservació del DNA original, hi ha els fòssils trobats l’any 1857 a la vall de Neander i que van servir per batejar el grup. Amb posterioritat, la mostra de neandertals en què ha estat possible purificar autèntic DNA mitocondrial s’ha anat ampliant, incloent-hi els jaciments de La Chapelle-aux-Saints a França, Engis a Bèlgica, Mezmaiskaia a Rússia i Teshik Tash a Uzbekistan. No obstant això, dos jaciments –Vindija a Croàcia i El Sidrón a Espanya– destaquen sobre la resta pel nivell de conservació del seu DNA, per la qual cosa han contribuït molt significativament a la descodificació d’aquesta molècula en els neandertals. En l’actualitat, la recuperació de DNA mitocondrial s’ha pogut estendre fins als avantpassats dels neandertals, com és el cas del jaciment Sima de los Huesos a Atapuerca, les restes humanes del qual, atribuïdes a Homo heidelbergensis, es remunten a prop de 500.000 anys.
Els primers resultats sobre DNA mitocondrial en neandertals van revelar l’existència de seqüències molt diferents de les del tipus humà actual (Krings et al., 1997). D’un total de 379 parells de bases, es van observar 27 diferències, que contrasten amb les escasses 5 o 8 diferències esperades per a una mostra a l’atzar entre els humans actuals. Això significava que la diferència general entre les seqüències dels neandertals i els humans moderns era tres vegades superior a la mitjana entre els humans actuals, però només la meitat de l’observada entre humans i ximpanzés. Si considerem el temps de divergència entre aquestes dues últimes espècies, això implicava una antiguitat notable per a la divergència entre la línia que va portar a l’home anatòmicament modern i la que va desembocar en els neandertals: entre fa 550.000 i 690.000 anys. El fet que les línies que van desembocar en els dos grups hagueren evolucionat independentment durant més de mig milió d’anys es va interpretar com una clara evidència que Homo neanderthalensis i Homo sapiens constituïen dues espècies diferents.
Aquest resultat posava punt final a un debat candent que es va desenvolupar a les acaballes del segle XX entre els partidaris del model multiregional, com Milford Wolpoff i Erik Trinkaus, i els partidaris del model «Arca de Noè», com Chris Stringer i Richard Klein, per a explicar l’origen de l’Homo sapiens. Segons Wolpoff i Trinkaus, després de la primera eixida d’Àfrica, les diferents poblacions d’Homo erectus van continuar mantenint un flux genètic elevat, de manera que diferents poblacions regionals haurien desenvolupat en paral·lel les característiques d’Homo sapiens. Enfront d’aquest model, la major part de paleontòlegs advocaven per un origen africà únic de la nostra espècie, previ a la dispersió per la resta de continents. Doncs bé, els resultats proporcionats per l’anàlisi del mitogenoma neandertal confirmaven aquest últim punt de vista i revelaven que neandertals i humans moderns constituïen espècies diferents.
El següent pas en la investigació va ser el Projecte Genoma Neandertal, abordat conjuntament pels equips de Richard E. Green, de la Universitat de Califòrnia, i Svante Pääbo, de l’Institut Max Plank a Alemanya. Aquesta vegada ja no es tractava de reconstruir el DNA mitocondrial sinó el genoma complet del neandertal, tal com s’havia fet amb el genoma humà anteriorment. Després d’anys d’investigació, en els quals de nou els jaciments de Vindija i El Sidrón van representar el paper més rellevant, el 2010 es va publicar un primer esborrany que incloïa aproximadament un 60 % de la seqüència gènica (Green et al., 2010) i arribava a conclusions que es van confirmar el 2013 amb la publicació del genoma complet. Sorprenentment, l’anàlisi del DNA neandertal va revelar la presència d’entre un 1 i un 4 % de seqüències neandertals en el genoma dels humans moderns euroasiàtics, però no en el genoma de les poblacions africanes actuals. Això volia dir que aquest limitat encreuament es degué produir en els primers moments de l’expansió de d’Homo sapiens fora de l’Àfrica, molt probablement quan ambdues poblacions es degueren trobar en el corredor de Palestina fa entre 40.000 i 60.000 anys. Els multiregionalistes i els partidaris de l’encreuament entre neandertals i humans moderns van acollir aquest resultat amb alegria, ja que interpretaven que, d’alguna manera, constituïa un aval per a les seues idees. En termes estrictes, aquesta dada implicava que, tècnicament, neandertals i sapiens no podrien considerar-se espècies diferents.
Per a la majoria de paleoantropòlegs, però, aquest resultat no modificava significativament la idea que neandertals i humans moderns representen en realitat espècies o quasiespècies diferents, amb històries evolutives divergents durant centenars de milers d’anys, sobretot si es té en compte que, després del seu contacte a Palestina, ambdues poblacions van tornar a coexistir a Europa durant més de 10.000 anys, sense que es tornaren a produir encreuaments efectius.
Però la revelació del genoma neandertal no va ser l’únic resultat sorprenent aportat per la paleogenòmica. L’anàlisi realitzada per l’equip de Pääbo d’un petit fragment de falange infantil procedent de nivells datats entre 48.000 i 30.000 anys de la cova de Deníssova, a la serralada d’Altai a Sibèria, va revelar una seqüència de nucleòtids diferents no sols de la dels humans actuals, sinó també dels neandertals (Krause et al., 2010). Els denisovans, com així van ser anomenats, havien habitat Àsia central i havien arribat a encreuar-se amb els neandertals i amb els humans moderns. De fet, entre les poblacions modernes, el genoma denisovà mostrava les màximes concomitàncies amb els habitants de Papua Nova Guinea. No obstant això, no hi havia rastre de gens denisovans en poblacions molt més pròximes, com els xinesos.
Ens trobem, per tant, davant d’un hominí anterior a neandertals i humans moderns, que va arribar a conviure-hi i fins i tot a encreuar-se. No obstant això, a part d’un parell de molars de grans dimensions i la falange infantil, no sabem res d’aquest misteriós hominí. L’estret parentiu amb els neandertals apuntaria poblacions relictes d’Homo heidelbergensis, però podria tractar-se d’hominins encara més arcaics, tal vegada poblacions relictes d’Homo erectus, que en un moment donat haurien arribat a encreuar-se amb humans moderns a Australàsia. En tot cas, la irrupció dels denisovans ha acabat trencant els motles en què s’havia basat la investigació paleoantropològica fins a l’actualitat.
REFERÈNCIES
Agustí, J., & Lordkipanidze, D. (2005). Los primeros pobladores de Europa. Barcelona: RBA.
Green, R. E., Krause, J., Briggs, A. W., Maricic, T., Stenzel, U., Kircher, M., ... Pääbo, S. (2010). A draft sequence of the Neandertal genome. Science, 328, 710–722. doi: 10.1126/science.1188021
Krause, J., Fu, Q., Good, J. F., Viola, B., Shunkov, M. V., Derevianko, A. P., & Pääbo, S. (2010). The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature, 464, 894–897. doi: 10.1038/nature08976
Krings, M., Stone, A., Schmitz, R. W., Krainitzki, H., Stoneking, & Pääbo, S. (1997). Neandertal DNA sequences and the origin of modern humans. Cell, 90, 19–30. doi: 10.1016/S0092-8674(00)80310-4
Lordkipanidze, D., Vekua, A., Ferring, R., Rightmire, G. P., Agustí, J., Kiladze, G., … Zollikofer, C. (2005). The earliest toothless hominin skull. Nature, 434, 717–718. doi: 10.1038/434717b
Lordkipanidze, D., Jashashvili, T., Vekua, A., Ponce de León, M., Zollikofer, C., Rightmire, … Nioradze, M. (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, 449, 305–310. doi: 10.1038/nature06134
Lordkipanidze, D., Ponce de León, M., Margvelashvili, A., Rak, Y., Rightmire, G. P., & Zollikofer, C. (2013). A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science, 342, 326–331. doi: 10.1126/science.1238484
Vekua, A., Lordkipanidze, D., Rightmire, G. P., Agustí, J., Ferring, R., Maisuradze, G., … Zollikofer, C. (2002). A new skull of early Homo from Dmanisi, Georgia. Science, 297, 85–89. doi: 10.1126/science.1072953
Villmoare, B., Kimbel, W. H., Seyoum, C., Campisano, C. J., DiMaggio, E., Rowan, J., … Reed, K. E. 2015. Early Homo at 2.8 Ma from Ledi Geraru, Afar, Ethiopia. Science, 347, 1352–1355. doi: 10.1126/science.aaa1343




