

Pròxima parada: Llenguatge
El viatge del gen FOXP2 a través del temps

Com va evolucionar el llenguatge en l’ésser humà? El registre fòssil no ofereix proves suficients per a reconstruir-ne l’evolució i els animals no parlen. Però a mesura que es van revelant els substrats moleculars i neurals del llenguatge, es poden abordar l’origen i funció de manera comparativa en altres espècies. Això és el que ocorre amb FOXP2, un gen estretament relacionat amb el llenguatge que és essencial per a l’aprenentatge en ratolins, ocells i fins i tot mosques. La comparació del paper del gen FOXP2 en humans i en altres animals està començant a revelar principis comuns que podrien haver proporcionat elements fonamentals per a l’evolució del llenguatge.
Paraules clau: parla, llenguatge, aprenentatge sensomotor, evo-devo, homologia profunda.

Figura 1. Què li semblaria més interessant a un científic extraterrestre que aterrara al planeta Terra? / Peter Schatton
El llenguatge és universal
Imagineu que sou un científic extraterrestre que estudia la conducta i que aterra al nostre planeta per primera vegada (figura 1). Què li sorprendria més? Les enormes balenes i les petites mosques de la fruita, aqueferades alimentant-se i reproduint-se de formes molt diferents? O li fascinarien els simis bípedes amb roba que ocupen hàbitats molt diferents de cap a cap del globus, semblen molt diversos i parlen entre ells amb cadenes de sons molt diferents, però que tot i això són part de la mateixa espècie, Homo sapiens? Si mirem més de prop, veurem que aquestes cadenes de sons diferents, que anomenem llengües, són, de fet, fonamentalment universals: totes les societats humanes tenen la seua pròpia llengua.
Encara que sonen distintes, totes les llengües comparteixen algunes similituds. Tots els idiomes s’estructuren de manera jeràrquica. Els elements fonamentals són els sons que els humans poden generar sense esforç amb la seua veu, com p, b, f, v, t, d, a, e i altres. Es combinen en un repertori més ampli de síl·labes amb què es poden compondre milers de paraules diferents. Les normes gramaticals, que també són parcialment semblants en tots els idiomes, permeten enfilar combinacions quasi infinites de paraules per a construir frases que poden expressar un nombre aparentment il·limitat d’accions i pensaments. El mateix s’aplica a les llengües de signes, en les quals un repertori limitat de gestos i moviments facials es combina per formar un repertori major de gestos que, al seu torn, gràcies a la seua gramàtica, formen oracions amb significat.
El nostre científic extraterrestre estudiós de la conducta podria pensar: «Quin sistema tan enginyós!» Però, com és que comparteixen principis comuns llengües tan diferents?
«A partir d’unes poques setmanes després del naixement, els nens poden distingir els sons de la parla d’altres sons»
Contribucions genètiques al llenguatge
El fet que tots els humans utilitzen el llenguatge i que totes les llengües compartesquen algunes característiques ha suggerit als lingüistes i biòlegs que deu existir un component genètic important. El component genètic del llenguatge, anomenat «instint del llenguatge» en el títol del llibre publicat el 1994 per Steven Pinker, deu estar relacionat amb la capacitat d’«aprendre» la llengua, ja que la composició genètica d’un nadó no afecta quina llengua concreta parlarà; un nadó nascut de pares de parla hispana però adoptat i criat per pares de parla francesa aprendrà francès, no espanyol. Llavors, com té lloc aquest procés d’aprenentatge? Primer, el cervell del nadó ha d’identificar quins sons són part de la llengua. Unes poques setmanes després del naixement, els nens poden destriar els sons de la parla d’altres sons i poden fins i tot relacionar una síl·laba en sentir-la amb la cara que ha produït el so (Kuhl i Meltzoff, 1982). Quan un nadó comença a balbotejar als pocs mesos d’edat, ha de modificar els seus balbotejos per ajustar-los a la llengua del seu entorn. Aquest procés continua durant els primers dos anys de vida. En paral·lel, el nadó aprèn –sense esforç i sense instrucció formal– el significat dels sons i les normes que governen la forma com es combinen. Fins i tot abans que els menuts puguen parlar bé, ja coneixen la diferència de significat quan els seus pares pregunten «Li has llevat el joguet a Maria?» o «Maria t’ha llevat el teu joguet?». A mesura que els nens creixen, els seus cervells també reaccionen de manera diferent davant d’oracions que són semànticament correctes o incorrectes («El meu oncle veurà la pel·lícula» enfront de «El meu oncle bufarà la pel·lícula»), i també diferencien entre oracions sintàcticament correctes i incorrectes («El meu oncle veurà/veient la pel·lícula») (Friederici, 2006).
«Si els nadons naixen amb un instint per a l’aprenentatge lingüístic, com es fixa això en els seus gens? O l’aprenentatge lingüístic només és un procés més d’aprenentatge?»
«Oh!», diu l’extraterrestre, «sembla que aquests nadons d’Homo sapiens naixen sent petites màquines d’aprendre llenguatge. Com ho fan?»
FOXP21
Si els nadons naixen amb un instint per a l’aprenentatge lingüístic, com es fixa això en els seus gens? Necessiten gens «especialitzats» que únicament són rellevants per a la parla o el llenguatge? O l’aprenentatge lingüístic només és un altre procés més d’aprenentatge, com aprendre a caminar, a anar amb bicicleta o matemàtiques? La segona opció semblava la més probable perquè les malalties genètiques que afecten el llenguatge també solen afectar altres dominis cognitius. Per això el diagnòstic d’un «trastorn específic del llenguatge» que no implicava altres anomalies morfològiques o mentals en una família britànica va sorprendre la comunitat científica. Sembla que l’àvia de la família coneguda com KE tenia una mutació espontània que va ser heretada per quatre dels seus cinc fills i filles i deu dels seus 24 néts i nétes. El patró hereditari és característic d’una mutació dominant en un dels cromosomes no sexuals (autosomes). En el cas de la família KE, els membres afectats tenen problemes per a coordinar la llengua i els músculs de la boca i produir sons de la parla. Per exemple, repetir cadenes polisil·làbiques sense sentit com «woogalamic» o «perplisteronk» és molt més complicat per a un membre de la família KE amb la mutació que per a un familiar no afectat (Watkins, Dronkers i Vargha-Khadem, 2002). A més, la comprensió gramatical també està afectada, encara que en menor grau que la producció de la parla. Per exemple, quan se’ls demanava que assenyalaren la imatge que correspon al text «La llebre que persegueix el cavall és roja», els membres de la família afectats cometien més errors (figura 2).
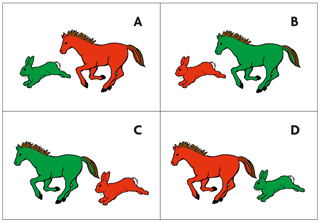
Figura 2. Imatge basada en el test TROG (test de recepció de gramàtica, Bishop, 1983). Els membres afectats de la família KE fallaven més quan se’ls demanava que assenyalaren la imatge adequada per al text: «La llebre que persegueix el cavall és roja.» / Adriana Schatton
Com que els problemes de comprensió i producció de llenguatge complex afecten l’educació i la interacció amb els altres, la mutació representa un handicap enorme per als membres afectats de la família. Al mateix temps, el cas va ser per a la comunitat científica una finestra oberta als fonaments moleculars de la producció i percepció lingüística, i va proporcionar l’esperança d’una possible intervenció clínica (Vargha-Khadem, Gadian, Copp i Mishkin, 2005). La cartografia genètica va revelar que tots els membres afectats de la família KE tenien una mutació en un gen del cromosoma 7 anomenat Forkhead box P2, o FOXP2 (Lai, Fisher, Hurst, Vargha-Khadem i Monaco, 2001). El curiós nom, forkhead box (“domini de cap de forquilla”), el va encunyar el 1989 un grup de científics que treballava amb la mosca de la fruita. Van identificar mutants amb «cap de forca» (forked head en llengua anglesa) i van connectar el fenotip amb una secció particular de l’ADN. Aquest domini existeix en més de quaranta gens organitzats en 19 famílies que reben noms que van des de FoxA fins a FoxS. Tots codifiquen proteïnes, anomenades «factors de transcripció», que regulen la funció d’altres gens. La part forkhead box del factor de transcripció s’uneix a regions reguladores específiques d’altres gens, denominats «gens diana», i incrementen o disminueixen la seua activitat. Molts factors de transcripció amb dominis forkhead estan relacionats amb la salut i la malaltia, però només la família de gens P s’ha relacionat amb la parla i el llenguatge.
El científic extraterrestre es rasca les seues pròpies «antenes en forma de forca» i es pregunta: «Si jo tinguera un gen FOXP2 podria parlar com Homo sapiens?»
«El descobriment del FOXP2 va obrir la porta a la possibilitat de respondre moltes preguntes sobre la connexió entre gens i llenguatge»
Un gen per a cada tret?
El descobriment del FOXP2 va obrir la porta a la possibilitat de respondre moltes preguntes sobre la connexió entre gens i llenguatge. Tenir un gen FOXP2 ens dóna el llenguatge? No. El FoxP2 existeix en moltes espècies que no parlen. La importància d’aquest gen per al desenvolupament pulmonar en ratolins ja s’havia descrit fins i tot abans que la seua mutació en la família KE es relacionara amb el llenguatge, i va mostrar que el FoxP2 no era ni un gen exclusivament humà ni exclusivament relacionat amb el cervell. Els gens exclusivament humans existeixen (Dennis et al., 2012) però FoxP2 no n’és un. FoxP2 existeix en els 274 vertebrats estudiats fins ara i es troba entre els gens la seqüència dels quals amb prou feines ha canviat en l’evolució dels vertebrats. Això suggereix que la funció del gen FoxP2 podria no haver canviat molt de peixos a humans, per la qual cosa siga quina siga la funció exacta d’aquest gen, hi ha una pressió evolutiva per conservar-ne la seqüència. Aquesta idea es veu reforçada pel descobriment que les mutacions del FOXP2 identificades en humans –a hores d’ara són 23 mutacions distintes i totes afecten la parla i el llenguatge– mai afecten les dues còpies del gen (al·lels), la qual cosa implica que quan les dues còpies han mutat, l’embrió no pot sobreviure. Això succeeix també en ratolins amb mutacions en ambdós al·lels del gen Foxp2: moren poques setmanes després del naixement, segons sembla per trastorns pulmonars. Les rates penades són una excepció a l’estesa conservació del FoxP2 en els vertebrats. Moltes espècies de rates penades tenen una seqüència FoxP2 molt divergent. Això és un descobriment enigmàtic que encara espera una explicació (revisat en Knörnschild, 2014).
«El component genètic del llenguatge ha d’estar relacionat amb la capacitat d’aprendre el llenguatge, ja que la composició genètica d’un nadó no afecta quina llengua concreta parlarà»
A l’extraterrestre, això li semblaria un poc confús. Les mutacions FOXP2 causen un dèficit en un tret exclusivament humà, però pràcticament la mateixa versió del FoxP2 existeix en tots els vertebrats?
De ratolins i homes
Just quan semblava que el FOXP2 no era al capdavall tan especial, Wolfgang Endard i els seus col·legues de l’Institut Max Planck d’Antropologia Evolutiva en Leipzig van informar que, a pesar que la seqüència de la proteïna FOXP2 en humans era en conjunt molt semblant a la FoxP2 en la resta de vertebrats, la variant humana havia canviat en la història (evolutiva) recent més que no s’esperava. A més, tots els humans tenen aquesta versió de FOXP2 específicament humana des de fa uns 200.000 anys, coincidint aproximadament amb l’aparició dels humans anatòmicament moderns, encara que el moment exacte és controvertit. La noció que la versió exclusivament humana d’aquesta proteïna va ser important per a l’evolució humana va ser rebuda amb escepticisme perquè la seqüència diferia de la versió dels ximpanzés només en dos dels 714 aminoàcids, els elements constitutius de les proteïnes. Podien només dos aminoàcids ser la raó per la qual nosaltres parlem i els ximpanzés no? D’altra banda, el descobriment d’una versió de FOXP2 específica en humans que semblava ser beneficiós mantenir i que, per tant, estava present en tota la població humana requeria més investigació. Per això, els científics de Leipzig van dissenyar un experiment enginyós: van afegir experimentalment el gen humà FOXP2 a ratolins i van comprovar si aquests eren diferents dels seus germans no manipulats (Enard et al., 2009). No va ser cap sorpresa, però els ratolins no van desenvolupar el llenguatge, i això provava que la versió humana de FOXP2 no pot, per si sola, desencadenar els canvis necessaris en el cervell i l’aparell vocal per a desenvolupar la parla i el llenguatge.
«Quan es va separar de sa mare les cries de ratolí que tenien la versió del FOXP2 humana, van fer crides d’auxili amb característiques bioacústiques diferents de les de les cries no manipulades»
Ateses les diferències entre els cervells i les estructures generadores de so d’un ratolí i un humà, era, per descomptat, molt poc probable que un únic gen poguera activar tal programa. No obstant això, l’experiment d’introduir el gen humà FOXP2 en un ratolí va ser informatiu; quan es va separar de sa mare les cries que tenien la versió del FOXP2 humana, van fer crides d’auxili amb característiques bioacústiques diferents de les de les cries no manipulades, la qual cosa representa una altra connexió entre FOXP2 i les vocalitzacions. Encara no es coneixen els substrats neurals per a aquests canvis de vocalització, però diversos experiments posteriors han mostrat que els cervells de ratolins exposats a la versió humanitzada de FOXP2 són diferents estructuralment i funcionalment dels cervells de ratolins amb Foxp2 de ratolí. La investigació posterior dels laboratoris d’Ann Graybiel, Simon Fisher i Wolfgang Enard va establir que els cervells de ratolins que s’havia alterat experimentalment perquè portaren una mutació equivalent a la de la família KE també mostren diferències amb els ratolins comuns. Curiosament, sembla que en els ratolins algunes de les conseqüències neurals i conductuals de tenir un al·lel Foxp2 amb una mutació semblant a la que produeix trastorns del llenguatge en humans són oposades a aquelles presents en ratolins amb la versió humanitzada del gen. Aquests últims també difereixen dels ratolins de ventrades normals en tasques de resolució de laberints. Quan els ratolins han de passar de trobar el camí basant-se en senyals visuals (com veure un estel) a trobar el camí girant a l’esquerra o a la dreta, aquells humanitzats amb FoxP mostren una transició més ràpida d’una estratègia a l’altra. Els autors suggereixen que l’aprenentatge lingüístic requereix un canvi semblant entre estils d’aprenentatge.
Junts, aquests resultats posen en relleu que petits canvis en la seqüència d’un gen molt conservat podrien canviar la funció neural, però que els trets de conducta o les adaptacions morfològiques únics no necessiten gens únics. Més aviat són combinacions concretes de grups de gens els que poden actuar en conjunt i provocar efectes «únics». De fet, des del descobriment original sobre la relació entre mutacions del FOXP2 i el trastorn específic del llenguatge, s’han identificat més gens implicats en diferents trastorns relacionats amb el llenguatge. És probable que alguns d’aquests gens actuen junt amb el FOXP2 en una xarxa molecular. Per exemple, un altre membre de la família genètica FoxP, FOXP1, també està relacionat amb dèficits del llenguatge i les proteïnes FoxP1 i FoxP2 poden interactuar directament (Graham i Fisher, 2015).
El científic extraterrestre crida: «Quina idea més bona introduir la versió humana d’un gen en ratolins!» I es pregunta per què es van produir canvis neurals i de comportament en aquests ratolins, encara que no començaren a parlar…
Formació de circuits versus funció de circuits
Va afectar la versió humana del FOXP2 el desenvolupament cerebral dels ratolins? O la manipulació genètica va afectar la funció cerebral després que es completara el desenvolupament? Les mutacions del FOXP2 en humans obstaculitzen el desenvolupament embrionari dels circuits cerebrals que els nens necessiten més avant per a l’aprenentatge lingüístic? O l’activitat del FOXP2 té un paper durant el mateix procés d’aprenentatge lingüístic? L’última pregunta no es pot contestar amb ratolins, perquè les seues vocalitzacions no són com el llenguatge humà, no s’aprenen imitant els sons d’adults de la seua mateixa espècie. Els nens que naixen sords no aprenen a parlar tan bé com els que senten, perquè l’aprenentatge de la parla depèn de la retroalimentació auditiva. Al contrari, els ratolins sords vocalitzen exactament igual que els ratolins amb audició, la qual cosa demostra que les vocalitzacions dels ratolins no s’aprenen per imitació.
A diferència dels ratolins, algunes espècies de balenes i de rates penades sí que modifiquen els sons que utilitzen per a comunicar-se a partir de l’experiència (Janik i Slater, 1997; Knörnschild, 2014). Aquests aprenents vocals són difícils de mantenir en el laboratori, per la qual cosa encara no s’han realitzat estudis de la funció genètica en rates penades i balenes. Moltes espècies d’aus canores també aprenen a produir el seu cant imitant adults de la seua espècie i aquest procés presenta moltes similituds conductuals i neurals amb l’aprenentatge de la parla (Bolhuis, Okanoya i Scharff, 2010) (figura 3). A més, ja es coneixen les estructures neurals subjacents a la producció i a l’aprenentatge del cant, i els genomes d’aproximadament cinquanta aus canores s’han seqüenciat recentment (Zhang et al., 2014), la qual cosa incrementa l’interès dels estudis neurogenètics comparatius. En aquest sentit, els investigadors del nostre grup a Berlín van comprovar que l’expressió de FoxP2 es modula durant el període d’aprenentatge del cant en una regió del cervell essencial per al dit aprenentatge (Haesler et al., 2004). A més, la reducció experimental dels nivells de FoxP2 en aquesta regió del cervell provoca una imitació del cant incompleta i inexacta en pinsans zebra (Haesler et al., 2007). Altres experiments nostres i d’altres investigadors han identificat factors que regulen l’expressió de FoxP2 i les conseqüències cel·lulars i conductuals de la seua acció (Condro i White, 2014; Wohlgemuth, Adam i Scharff, 2014). Aquests i altres estudis han establit amb fermesa que FoxP2 té dues funcions, una en el desenvolupament neural i una altra «en directe» en circuits neurals ja madurs (Murugan, Harward, Scharff i Mooney, 2013).
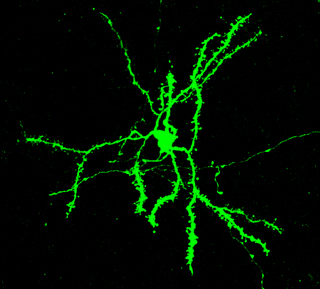
Figura 4. En ocells canors i en mamífers, la proteïna FoxP2 s’expressa en el que es coneix com a neurones espinoses mitjanes del cos estriat, una regió important per a convertir informació sensorial en conductes motores. Es coneix aquestes neurones com espinoses perquè tenen «espines», punts en què es reben senyals d’altres neurones. / Jennifer Kosubek
L’investigador extraterrestre recapitula: «Les aus canores aprenen a cantar de manera semblant a com els humans aprenen a parlar, i els dos necessiten FoxP2 per a fer-ho bé. Com pot ser això?»
Temes comuns
El fet que FoxP2 siga important tant per a la parla humana com per al cant de les aus, dos comportaments molt semblants, i també per a l’aprenentatge motor en els ratolins, és molt interessant des d’una perspectiva evolutiva. El que tenen en comú és que aquest factor de transcripció actua en circuits neurals que són rellevants per a la conversió d’estímuls sensorials (auditius per a humans i aus, visuals i tàctils per als ratolins) en accions motores (la parla en humans, el cant en les aus i el moviment en ratolins). Els circuits neurals centrals per a tots aquests comportaments són els que es coneixen com a ganglis basals, que són homòlegs en aus, ratolins i humans. «Homòleg» en aquest context significa que aquestes estructures semblen derivar d’estructures que ja existien en l’avantpassat comú d’aus i mamífers fa 300 milions d’anys. La forma i la funció dels ganglis basals són sorprenentment semblants en aus i mamífers. Una teoria popular, avalada per molts resultats empírics, defensa que l’aprenentatge sensomotor en aquest circuit depèn d’un sistema neuroquímic de recompensa (per exemple, la dopamina) (figura 4). Les proves experimentals en ratolins i aus suggereixen que FoxP2 i la seua xarxa molecular associada, incloent-hi les altres dues proteïnes expressades neuronalment, FoxP1 i FoxP4, interactuen amb el senyal de la dopamina i regulen la potència de les connexions concretes entre conjunts determinats de neurones, afinant la integració sensomotora (Wohlgemuth et al., 2014).
«El fet que FoxP2 siga important tant per a la parla humana com per al cant de les aus és molt interessant des d’una perspectiva evolutiva»
«Ah, comence a veure el fil comú que uneix tota aquesta història», dirà esperançat l’extraterrestre. FOXP2, junt amb altres molècules, sembla estar activa en parts del cervell que converteixen l’experiència sensorial en acció motora. Podria ajudar un nadó a modificar els seus balbucejos per a crear paraules, els ocells joves a ajustar els seus sons per a sonar de manera pareguda a la cançó de son pare i un ratolí a aprendre a usar el seu cos amb eficàcia per a aconseguir menjar i evitar ser menjat.
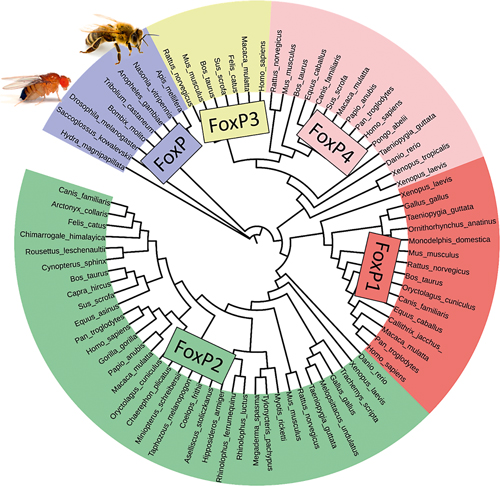
Figura 5. El gen FoxP dels invertebrats va donar lloc a través de la duplicació genètica a quatre gens FoxP diferents. En els vertebrats, FoxP1, 2 i 4 són importants per al desenvolupament del cervell i la conducta i també compleixen una funció clau per a altres òrgans. FoxP3 actua en el sistema immune. / Ezequiel Mendoza
Retrocedir en el temps
Convertir l’experiència sensorial en accions motores és un requisit per a la supervivència que no és exclusiu dels vertebrats. Si la proteïna FoxP2 és important per a aquesta funció, existia ja abans que el llinatge dels vertebrats apareguera? En efecte, el locus del gen FoxP2 que va donar lloc als quatre gens FoxP diferents en els vertebrats és molt antic. Sembla que aquests gens van aparèixer alhora que la pluricel·lularitat. Els coanoflagelats unicel·lulars no tenen el gen però les esponges i altres animals sense simetria bilateral sí que en tenen (Shimeld, Degnan i Luke, 2010) (figura 5). En les esponges, FoxP realitza funcions d’adherència cel·lular (sobre un plat de plàstic) i en la formació d’agregats en capes semblants a l’epiteli. En el pulmó del ratolí, FoxP1 i FoxP4 participen en el desenvolupament de l’epiteli pulmonar (Li et al., 2012). Seran necessaris estudis més profunds per comprovar si es tracta d’una «coincidència epitelial» o una homologia més profunda. En el llinatge Bilateria, animals amb simetria bilateral, també hi ha un sol gen FoxP, amb dues versions distintes anomenades isoformes. FoxP s’expressa en el sistema nerviós embrionari i adult de la mosca de la fruita, el que suggereix que podria tenir una funció similar a la que té en vertebrats (Santos, Athanasiadis, Leitão, DuPasquier i Sucena, 2011).
Per a saber si FoxP podria tenir funcions semblants en invertebrats i en vertebrats, el nostre grup i altres hem començat a estudiar-ne el patró d’expressió i la funció en mosques de la fruita (Drosophila melanogaster) i abelles (Apis mellifera) (Kiya, Itoh i Kubo, 2008; DasGupta, Ferreira i Miesenböck, 2014; Lawton, Wassmer i Deitcher, 2014; Mendoza et al., 2014) (figura 6). Com sol ocórrer en les etapes inicials de la investigació científica d’una qüestió nova, alguns resultats coincideixen, mentre que d’altres discrepen. Quant als resultats coincidents, totes les mosques mutants FoxP mostren dèficits conductuals interessants que a primera vista no són incompatibles entre ells i també podrien estar relacionats conceptualment amb els dèficits conductuals observats en els vertebrats amb dosis reduïdes de FoxP2. Els autors d’un estudi van observar una reducció en les conductes de festeig (Lawton et al., 2014), en un altre estudi les mosques van aprendre a utilitzar senyals visuals per a evitar un càstig però no van poder utilitzar informació somatosensorial per a la mateixa tasca (Mendoza et al., 2014), i, en un tercer estudi, les mosques mutants van tenir problemes per a prendre decisions utilitzant senyals olfactius en diferents concentracions i el càstig com a incentiu d’aprenentatge (DasGupta et al., 2014). En aquest últim experiment, el fenotip es va poder atribuir a un petit grup determinat de neurones que anteriorment s’havien descrit com importants per a la presa de decisions i que ara es demostrava que també expressaven FoxP.
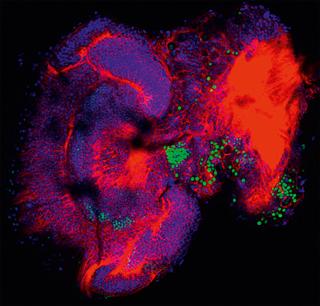
Figura 6. En el cervell d’una larva d’abella, FoxP s’expressa clarament en diferents conjunts de neurones (verd), indicant la importància del gen per al desenvolupament.
Hi ha discrepàncies pel que fa al patró d’expressió neural de FoxP en Drosophila, fins i tot quan es van utilitzar enfocaments pràcticament idèntics per a visualitzar-la. Utilitzant una proteïna fluorescent verd per detectar l’activitat promotora de FoxP, Lawton et al. (2014) van observar expressió en el que es coneix com a complex central, el qual podria ser funcionalment homòleg als ganglis basals de vertebrats segons algunes hipòtesis (Strausfeld i Hirth, 2013). Al contrari, utilitzant una seqüència promotora lleugerament distinta, DasGupta et al. (2014) no van trobar expressió de FoxP en el complex central, sinó en la regió nuclear dels cossos pedunculats (mushroom bodies), un lloc d’integració multimodal en el cervell de la mosca que és important per a l’aprenentatge i la memòria. Aquestes diferències també podrien explicar el fet que un estudi va indicar que les conductes motores (caminar) eren normals (DasGupta et al., 2014), mentre que en altres estudis (Lawton et al., 2014) les conductes motores es van deteriorar. No cal dir que, en les condicions experimentals, el diable està en els detalls; per la qual cosa, per refinar els resultats encara confusos de l’enfocament comparat, seran necessaris més estudis amb noves aproximacions que determinen els llocs d’expressió de l’ARNm i la proteïna FoxP en el sistema nerviós adult i en desenvolupament de mosques i d’abelles.

Figura 7. El nostre científic extraterrestre ha après molt sobre un gen i el seu viatge a través del temps. / Peter Schatton
Potser el nostre observador extraterrestre decidesca quedar-se un poc més en aquest planeta, arromangar-se i unir-se als científics terrestres per tractar de descobrir si FoxP és o no un candidat a «homologia profunda» que realitza funcions semblants en neurones d’espècies que van des de les mosques a Homo sapiens. O potser el nostre visitant espacial estiga cansat de tanta xarrameca per tot el planeta i desitge tornar-se a l’espai per pensar en silenci (figura 7).
És el llenguatge més que la suma de les seues parts?
Acompanyar FoxP en el seu viatge a través de l’evolució és interessant perquè apunta cap a un dels elements essencials de l’aprenentatge lingüístic: la conversió d’informació sensorial en accions motores. Però el llenguatge és, per descomptat, més que això. El llenguatge permet a la gent dir la veritat o explicar mentides, parlar sobre els esdeveniments d’ahir i els plans de demà, de conceptes abstractes com la lògica i la moral, de sentiments com l’alegria o la tristesa. No s’ha relacionat cap gen directament amb el procés pel qual els sentiments i pensaments s’externalitzen mitjançant la parla. Tal vegada un equip de genetistes i lingüistes extraterrestres que aterren a la Terra escriurà el pròxim capítol en el nostre camí cap a la comprensió de l’evolució del llenguatge.
- Per convenció, el gen humà s’escriu en majúscules, FOXP2; Foxp2 es refereix a la versió del gen en ratolins, i FoxP2 s’utilitza per a totes les altres espècies. Els gens s’escriuen en cursiva, les proteïnes, no. (Tornar al text)
REFERÈNCIES
Bolhuis, J. J., Okanoya, K., & Scharff, C. (2010). Twitter evolution: Converging mechanisms in birdsong and human speech. Nature Reviews Neuroscience, 11, 747–759. doi: 10.1038/nrn2931
Condro, M. C., & White, S. A. (2014). Recent advances in the genetics of vocal learning. Comparative Cognition & Behavior Reviews, 9, 75–98. doi: 10.3819/CCB.2014.90003
DasGupta, S., Ferreira, C. H., & Miesenböck, G. (2014). FoxP influences the speed and accuracy of a perceptual decision in Drosophila. Science, 344, 901–904. doi: 10.1126/science.1252114
Dennis, M. Y., Nuttle, X., Sudmant, P. H., Antonacci, F., Graves, T. A., Nefedov, M., … Eichler, E. E. (2012). Human-specific evolution of novel SRGAP2 genes by incomplete segmental duplication. Cell, 149, 912–922. doi: 10.1016/j.cell.2012.03.033
Enard, W., Gehre, S., Hammerschmidt, K., Hölter, S. M., Blass, T., Somel, M., … Pääbo, S. (2009). A humanized version of Foxp2 affects cortico-basal ganglia circuits in mice. Cell, 137, 961–971. doi: 10.1016/j.cell.2009.03.041
Friederici, A. D. (2006). The neural basis of language development and its impairment. Neuron, 52, 941–952. doi: 10.1016/j.neuron.2006.12.002
Graham, S. A., & Fisher, S. E. (2015). Understanding language from a genomic perspective. Annual Review of Genetics, 49, 131–160. doi: 10.1146/annurev-genet-120213-092236
Haesler, S., Rochefort, C., Georgi, B., Licznerski, P., Osten, P., & Scharff, C. (2007). Incomplete and inaccurate vocal imitation after knockdown of FoxP2 in songbird basal ganglia nucleus area X. PLoS Biology, 5, e321. doi: 10.1371/journal.pbio.0050321
Haesler, S., Wada, K., Nshdejan, A., Morrisey, E. E., Lints, T., Jarvis, E. D., & Scharff, C. (2004). FoxP2 expression in avian vocal learners and non-learners. The Journal of Neuroscience, 24, 3164–3175. doi: 10.1523/JNEUROSCI.4369-03-2004
Janik, V. M., & Slater, P. J. B. (1997). Vocal learning in mammals. En P. Slater, J. Rosenblatt, Ch. Snowdon, & M. Milinski (Eds.), Advances in the study of behavior. Vol. 24 (pp. 59–99). San Diego, CA: Academic Press.
Kiya, T., Itoh, Y., & Kubo, T. (2008). Expression analysis of the FoxP homologue in the brain of the honeybee, Apis mellifera. Insect Molecular Biology, 17, 53–60. doi: 10.1111/j.1365-2583.2008.00775.x
Knörnschild, M. (2014). Vocal production learning in bats. Current Opinion in Neurobiology, 28, 80–85. doi: 10.1016/j.conb.2014.06.014
Kuhl, P., & Meltzoff, A. (1982). The bimodal perception of speech in infancy. Science, 218, 1138–1141. doi: 10.1126/science.7146899
Lai, C. S. L., Fisher, S. E., Hurst, J. A., Vargha-Khadem, F., & Monaco, A. P. (2001). A forkhead-domain gene is mutated in a severe speech and language disorder. Nature, 413, 519–523. doi: 10.1038/35097076
Lawton, K. J., Wassmer, T. L., & Deitcher, D. L. (2014). Conserved role of Drosophila melanogaster FoxP in motor coordination and courtship song. Behavioural Brain Research, 268, 213–221. doi: 10.1016/j.bbr.2014/04.009
Li, S., Wang, Y., Zhang, Y., Lu, M. M., DeMayo, F. J., Dekker, J. D., Tucker, P. W., & Morrisey, E. E. (2012). Foxp1/4 control epithelial cell fate during lung development and regeneration through regulation of anterior gradient 2. Development, 139, 2500–2509. doi: 10.1242/dev.079699
Mendoza, E., Colomb, J., Rybak, J., Pflüger, H.-J., Zars, T., & Scharff, C. (2014). Drosophila FoxP mutants are deficient in operant self-learning. Plos One, 9(6), e100648. doi: 10.1371/journal.pone.0100648
Murugan, M., Harward, S., Scharff, C., & Mooney, R. (2013). Diminished FoxP2 levels affect dopaminergic modulation of corticostriatal signaling important to song variability. Neuron, 80(6), 1464–1476. doi: 10.1016/j.neuron.2013.1009.1021
Santos, M. E., Athanasiadis, A., Leitão, A. B., DuPasquier, L., & Sucena, É. (2011). Alternative splicing and gene duplication in the evolution of the FoxP gene subfamily. Molecular Biology and Evolution, 28, 237–247. doi: 10.1093/molbev/msq182
Shimeld, S. M., Degnan, B., & Luke, G. N. (2010). Evolutionary genomics of the Fox genes: Origin of gene families and the ancestry of gene clusters. Genomics, 95, 256–260. doi: 10.1016/j.ygeno.2009.08.002
Strausfeld, N. J., & Hirth, F. (2013). Deep homology of arthropod central complex and vertebrate basal ganglia. Science, 340, 157–161. doi: 10.1126/science.1231828
Vargha-Khadem, F., Gadian, D. G., Copp, A., & Mishkin, M. (2005). FOXP2 and the neuroanatomy of speech and language. Nature Reviews Neuroscience, 6, 131–138. doi: 10.1038/nrn1605
Watkins, K. E., Dronkers, N. F., & Vargha-Khadem, F. (2002). Behavioural analysis of an inherited speech and language disorder: Comparison with acquired aphasia. Brain, 125, 452–464. doi: 10.1093/brain/awf058
Wohlgemuth, S., Adam, I., & Scharff, C. (2014). FoxP2 in songbirds. Current Opinion in Neurobiology, 28, 86–93. doi: 10.1016/j.conb.2014.06.009
Zhang, G., Li, B., Li, C., Gilbert, M., Jarvis, E., Wang, J. & Consortium, T. A. G. (2014). Comparative genomic data of the Avian Phylogenomics Project. GigaScience, 3, 26. doi: 10.1186/2047-217X-3-26
AGRAÏMENTS:
Donem les gràcies a Artur i Anna Zychlinsky pels seus útils comentaris editorials, a Kate Watkins per subministrar les paraules sense sentit per a avaluar l’execució de la parla i a Peter Schatton per prestar el seu talent artístic per crear les figures 1 i 7, i a Ezequiel Mendoza per modificar la figura 5. Demanem disculpes als autors els descobriments dels quals no apareixen en les referències a causa de constriccions d’espai. La investigació realitzada en el laboratori de les autores ha rebut finançament de l’Excellence Cluster Neurocure i l’SFB 665 «Developmental Disturbances of the Nervous System».




