

Reflexions sobre l’origen de la vida
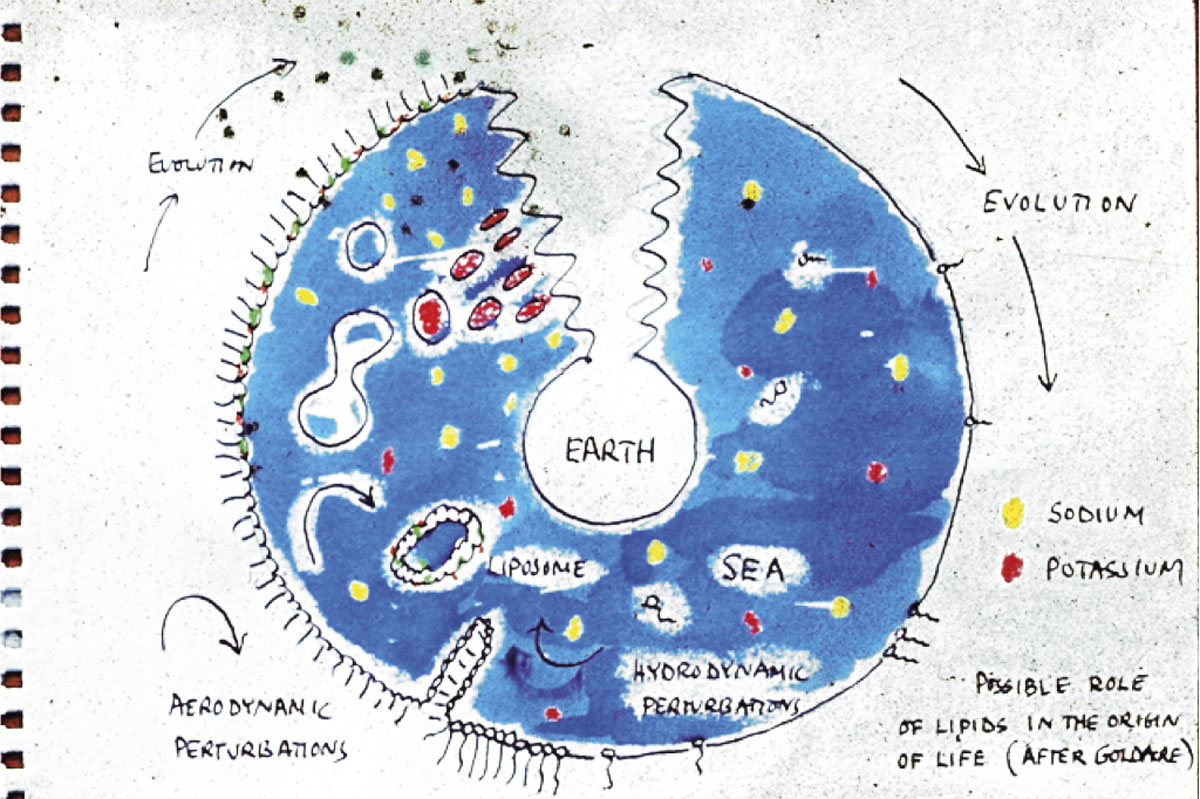
En aquest article argumentem que el problema de l’origen de la vida no es pot explicar apel·lant exclusivament als mecanismes de l’evolució darwiniana, com un gran nombre d’experts tendeix a pensar, sinó que requereix un profund canvi de perspectiva. En aquesta línia, destaquem que la selecció natural, per a operar com a motor de diversificació (i, indirectament, de potencial increment de la complexitat), implica un conjunt de condicions que, de fet, la facen possible: més en concret, organitzacions químiques automantingudes i auto(re)productives amb un espai fenotípic (és a dir, un ventall de funcions) bastant ampli. Així doncs, com a alternativa als plantejaments majoritaris en el camp de l’origen de vida, basats en poblacions moleculars (habitualment d’RNA) subjectes a evolució darwiniana, proposem una extensió del paradigma «autoorganitzatiu» cap a un d’«auto(re)productiu», que aplegue adequadament l’especificitat del fenomen biològic (en particular, la seua dimensió cel·lular i metabòlica), i que tindria rellevància abans, durant i després que la selecció natural començara a operar.
Paraules clau: autoorganització, protocèl·lules, metabolisme, diversitat fenotípica/funcional, evolució oberta.
Molts científics i filòsofs comparteixen actualment la idea que l’origen dels éssers vius a partir de la matèria inerta, tot i ser una problemàtica complicada i encara sense acabar de tancar, es troba, diguem-ne, «en via de solució». Òbviament falta un gran nombre de «detalls» per resoldre, que sens dubte poden ser difícils qüestions per als especialistes del camp; però, des d’una perspectiva global, s’assumeix que podem arribar a entendre aquesta transició fonamental, gràcies als resultats dels experiments de Stanley Miller i les diverses seqüeles que han tingut (és a dir, al camp de la «química prebiòtica» –que ha assolit la síntesi de diversos compostos rellevants per a la vida) i a la teoria de l’evolució, aplicada a les primeres molècules replicatives (nucleòtids d’RNA o anàlegs). No obstant això, algunes veus crítiques han posat en dubte aquesta visió tan optimista. Per exemple, recentment en el seu llibre Mind and Cosmos. Why the Materialist Neo-Darwinian Conception of Nature is Almost Certainly False, el filòsof Thomas Nagel (2012) sosté que les ciències biològiques contemporànies són incapaces d’explicar l’origen de la vida, o l’origen de la ment. Més concretament, Nagel afirma que cap teoria científica pot explicar satisfactòriament –el que per a ell significa una explicació reduccionista a les ciències físiques– com és que van aparèixer fenòmens tan complexos a partir de l’evolució del món fisicoquímic. Nagel en el seu llibre tendeix a identificar el sentit explicatiu del terme reduccionisme com a equivalent a materialisme reduccionista, perquè no considera que les teories «emergentistes» oferesquen una explicació alternativa d’aquest tipus de sistemes d’alta complexitat o de les seues propietats respectives, biològiques i cognitives. Com ell mateix diu: «Que aquests elements purament físics, quan són combinats d’una certa manera, hagen de produir necessàriament un estat del tot que no es constitueix a partir de les propietats i les relacions atribuïbles a les seues parts físiques, continua sonant-me a màgia».
«Una consideració detinguda sobre les diferències entre la matèria viva i la no viva ha de portar-nos a reflexionar més sobre les dificultats que tanca el problema de l’origen de la vida»
I és que, efectivament, una consideració detinguda sobre les diferències entre la matèria viva i la no viva ha de dur-nos a reflexionar més sobre les dificultats que conté el problema de l’origen de la vida. Explicar la transició entre el món fisicoquímic i el biològic és un repte d’enormes dimensions, principalment pel contrast tan profund existent entre ambdós dominis avui dia. Qualsevol ésser viu conegut, fins i tot unicel·lular, és ordres de magnitud més complex (en composició, diversitat d’interaccions, comportament dinàmic) que els distints sistemes moleculars dels quals s’ha postulat que poguera provenir. Quan analitzem els components, les transformacions químiques, els mecanismes de control que subjauen a les propietats de les cèl·lules més senzilles sobre el nostre planeta (procariotes amb genomes molt reduïts), és impossible entendre, d’una sola vegada, tot el que està ocorrent en el seu interior i en la seua relació amb l’entorn. Incomptables reaccions de síntesi (bio)química, juntament amb processos de transport, transducció, reconeixement, senyalització… ocorren cada segon en la forma necessària perquè l’organisme es mantinga, s’adapte i, potencialment, es reproduesca. Això és possible gràcies a una bateria de mecanismes moleculars que els mateixos éssers vius han generat (en particular, mecanismes metabòlics, de síntesi autònoma i control regulatiu [Ruiz-Mirazo i Moreno, 2012]), que els fan especialment robustos, tot i ser sistemes oberts i allunyats de l’equilibri termodinàmic. Cap altre tipus de sistema material conegut, natural o artificial, exhibeix, ni de lluny, nivells comparables de complexitat, tant en un sentit estructural com organitzatiu o funcional.
El contrast entre ambdós mons resulta molt evident si examinem cada ésser viu individualment, però també quan considerem el fenomen biològic des d’una perspectiva temporal més àmplia. A escales de temps molt majors que les fisiològiques, la vida se’ns manifesta com a evolució, és a dir, com un encadenament històric entre organitzacions o entitats efímeres, que tot i això transmeten a les seues successores els seus trets específics. És veritat que, si ampliem el marc prou, tot (fins i tot un àtom, una llei física) pot entendre’s com a fruit d’un procés de transformació, d’evolució temporal. No obstant això, una vegada que les peces i les regles d’interacció bàsiques van quedar establides, la matèria ha implementat modes d’evolució a llarg termini sorprenentment diferents. D’una banda, la matèria inanimada coneguda s’ha regit de manera estricta per les lleis fonamentals que caracteritzen el nostre univers físic en expansió, i ha generat estels, planetes, discos d’asteroides, pols interestel·lar, galàxies i forats negres. D’una altra, la matèria que ha integrat (encara que siga només localment i temporalment) els sistemes biològics ha contribuït al desenvolupament de mecanismes de control sobre la seua variació molt més sofisticats i indirectes, a pesar de ser plenament congruents amb les lleis fisicoquímiques.
Aquests mecanismes moleculars de control de la variació, partint curiosament de condicions d’estabilitat precària (estats estacionaris allunyats de l’equilibri termodinàmic), es van anar fixant d’una manera cada vegada més consistent en diverses poblacions de protoorganismes, als quals van conferir major robustesa i adaptabilitat metabòlica. Així, van aconseguir transformar-se, al llarg del procés d’origen de vida, en mòduls cada vegada més complexos i, alhora, de major perdurabilitat, ordres de magnitud major que el temps de vida mitjà d’aquells protoorganismes dels quals, de fet, provenien. Temps després, certs polímers proclius a còpia i conservació de la seua seqüència, esdevingueren «registres moleculars» (Pattee, 1969) parcials, però molt fiables, de la complexitat estructural i organitzativa de les cèl·lules en què es van anar allotjant i dins de les quals han anat prenent cos i sentit des de llavors fins avui. Va ser justament l’emergència i funcionalitat d’aquests «components registre», amb capacitat de replicació i transferència fiable a altres sistemes cel·lulars semblants, el que va permetre que la vida reeixira a establir-se com un fenomen sostenible a llarg termini sobre la superfície de la Terra, gràcies a una forma d’evolució de la matèria que havia estat inaccessible fins a aquell moment: l’evolució darwiniana (Lewontin, 1970) o evolució oberta (Ruiz-Mirazo i Moreno, 2012). Per sorprenent que puga semblar, actualment disposem d’evidència molt sòlida que aquest mode de canviar i reinventar-se que caracteritza els éssers vius, basat en poblacions de metabolismes genèticament instruïts, porta en marxa, almenys sobre el nostre planeta, uns quants milers de milions d’anys.
La selecció natural no explica l’origen
La importància del procés evolutiu biològic i la seua formidable profunditat temporal, com ens va mostrar Darwin –i com tota la biologia posterior a ell ha apuntalat– ha portat a atorgar a aquests mecanismes de control de la varietat molecular (els mecanismes genètics, hereditaris) un paper fonamental en el desplegament del domini dels éssers vius sobre la Terra. Això, juntament amb la falta d’una teoria adequada sobre l’organització biològica (sobre la qual tornarem de seguida), ha generat un important desequilibri que queda plasmat en concepcions probablement errònies –o si més no fortament descompensades– sobre el problema de l’origen de vida. Ens referim, en particular, a l’assumpció que la teoria de l’evolució (i, més específicament, el mecanisme de «selecció natural» aplicat a poblacions de molècules autoreplicants, com els RNA) siga la clau per a resoldre la transició entre el domini de les coses inertes i el dels éssers vius. Aquesta assumpció, àmpliament acceptada en cercles científics des de fa unes quantes dècades (Eigen i Schuster, 1979; Orgel, 1992; Szostak, 2012), és paradoxalment contrària a la intuïció del mateix Darwin, el qual, a pesar de tenir una visió de l’origen molt avançada (semblant en molts aspectes a la que es va forjar un segle més tard), considerava la ciència del seu temps, incloent-hi la teoria sobre l’evolució que ell mateix estava impulsant en aquells moments, limitada per a poder encarar aquesta problemàtica (Peretó, Bada i Lazcano, 2009).
«Ha estat una interpretació simplista del llegat de Darwin el que ha portat a considerar la selecció natural com la principal –per a alguns fins i tot l’única– força o guia en el procés d’emergència de vida»
En realitat ha estat una interpretació simplista del llegat de Darwin el que ha portat a considerar la selecció natural com la principal –per a alguns fins i tot l’única– «força» o «guia» en el procés d’emergència de vida. No obstant això, com argumentem amb més detall en Moreno i Ruiz-Mirazo (2009), basant-nos en autors com Lewontin (1970) o Wicken (1987), entre d’altres, l’establiment d’un mode d’evolució darwinià, que realment opere per selecció natural, requereix com a condició de partida un espai fenotípic bastant ampli, és a dir, un potencial de variació i diversificació funcional que resulta difícilment abastable per a formes d’organització material per sota del llindar de complexitat assimilable al de protocèl·lules amb metabolisme propi. En altres paraules, el mecanisme d’evolució per selecció natural no serveix per a explicar l’origen de la vida perquè, en el fons, la selecció natural implica ja l’existència d’organismes, o «protoorganismes» amb un fenotip (o un conjunt de funcions atribuïbles als seus components) bastant ric. Llavors, on buscar principis o mecanismes que expliquen com, en les condicions de la Terra primitiva, s’haja pogut generar un procés sostingut de canvis que portara a l’aparició de la vida? Hi ha alguna llei o principi en el món físic que permet concebre l’aparició de la vida com una necessitat o hem de resignar-nos a pensar, com ens deia Monod (1970), que l’origen de la vida és tan improbable, tan fruit de l’atzar, que es tracta pràcticament d’un miracle? Com pot la matèria originar una cosa que sembla tan profundament diferent de les seues propietats inicials?
En què consisteix el canvi de perspectiva que proposem?
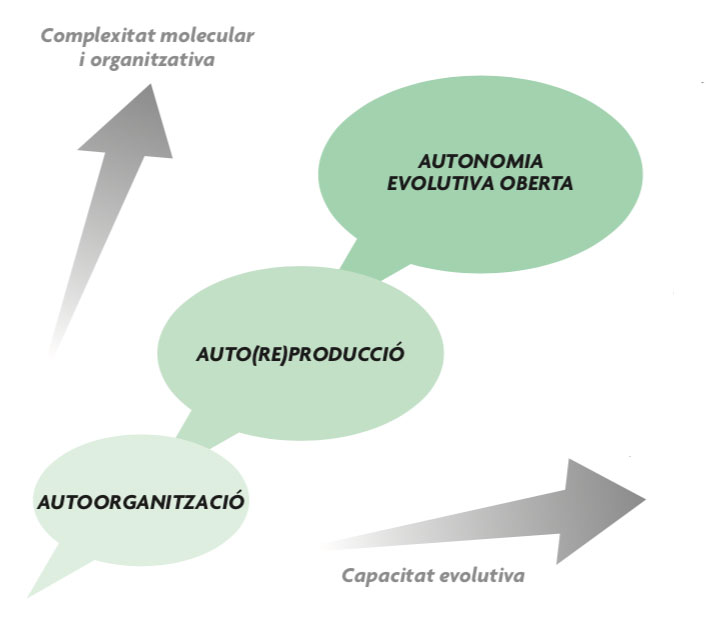
Una premissa de la qual podem partir, amb garanties de consens ple en la comunitat científica, és que la naturalesa no fa salts en el buit i, per tant, s’ha de plantejar una continuïtat entre la matèria no vivent i la vida. És a dir, ha d’haver-hi algun tipus d’explicació progressiva, en etapes de creixent complexitat (vegeu figura 1), de la transició cap a la vida, encara que en el límit ens podem veure abocats a una narrativa històrica, en gran manera contingent, del procés. Com Fry (2000) ha assenyalat, el problema fonamental de l’origen de la vida radica en la tensió entre el principi de continuïtat i la dificultat d’explicar les diferències òbvies entre la no-vida i la matèria viva. Si l’origen de la vida és una qüestió científica legítima (i pensem que ho és), hom ha de buscar una teoria que servesca de pont entre la física i la biologia, i demostrar experimentalment que aquest pont (almenys una part important d’aquest) es pot reconstruir de novo. Al capdavall, els éssers vius estan fets dels mateixos elements materials que els sistemes no vius, així doncs, no sembla desgavellat tractar d’aclarir el vincle que degué establir-se entre els uns i els altres.
«La naturalesa no fa salts en el buit i, per tant, s’ha de plantejar un continu entre la matèria no vivent i la vida»
Des de la perspectiva de les ciències físiques, explicar l’origen de la vida és una tasca molt difícil perquè com més complex és un sistema, menys probable és que aparega i que persistesca. En primera aproximació, sembla més o menys fàcil d’entendre que agregats materials simples puguen generar espontàniament estructures compostes relativament estables (molècules complexes, macromolècules o agregats supramoleculars) en virtut dels diferents tipus i graus de força que es donen en la naturalesa (Simon, 1962): efectivament, com a resultat d’aquestes interaccions físiques de distinta magnitud, és previsible la formació d’estructures més complexes (amb propietats que en molts casos es podrien considerar «emergents», no presents en les seues parts separades –com per exemple, la superconductivitat, l’afinitat química, o la topologia tancada d’una vesícula lipídica). Però a mesura que la complexitat de les estructures creix, mantenir-les esdevé un problema: el soroll tèrmic augmenta la fragilitat i, d’altra banda, la coincidència o la coordinació de molts processos altament específics es converteix en un fet cada vegada més improbable. A més, la complexitat de la vida no sols resideix en l’estructura dels seus components, sinó en una intricada xarxa d’interaccions dinàmiques que s’estableixen en condicions allunyades de l’equilibri termodinàmic. Dit d’una altra manera, estem obligats a encarar el problema en termes organitzatius. Els sistemes biològics, de fet, fan ús de les propietats autoorganitzatives de la matèria en molt diverses formes (Karsenti, 2008), unes més allunyades de l’equilibri que altres. I gràcies als avenços que es van donar durant la segona part del segle xx en química supramolecular i en termodinàmica de processos irreversibles, disposem d’un marc adequat per a entendre la formació d’organitzacions dinàmiques relativament complexes, que podrien arribar a combinar distints agregats supramoleculars i patrons d’ordre de tipus «estructura dissipativa».
No obstant això, la vida va més enllà: no sols organitza components materials preexistents, sinó que genera i regenera contínuament la majoria d’aquests i, alhora, passa a controlar bona part de les condicions en què aquesta organització dinàmica té lloc. És a dir, tot sistema biològic construeix una organització dinàmica a través de processos de síntesi i transformació de components, incloent-hi aquells que procuren les seues pròpies condicions de viabilitat i manteniment en el temps. Les implicacions d’aquest pas des de l’autoorganització a l’autoproducció són múltiples, però es poden resumir en dos (Ruiz-Mirazo i Moreno, 2012): (i) un tipus de cohesió interna i robustesa dinàmica molt major, basada en la «integració funcional» dels components del sistema; i (ii) un incipient control sobre les condicions de contorn sota les quals opera aquest sistema, és a dir, l’aparició de l’«agencialitat». Tot això està relacionat, en suma, amb l’emergència dels sistemes metabòlics: una problemàtica que posa en evidència que si aspirem a elaborar una teoria general, sòlida i congruent, sobre l’organització biològica, aquesta ha de prendre en consideració allò que han aportat les ciències de la complexitat i el paradigma de l’autoorganització, però, sens dubte, sobrepassant-lo, acostant-se d’una manera més específica a la fenomenologia de la vida (Keller, 2007).
«Des de la perspectiva de les ciències físiques, explicar l’origen de la vida és una tasca molt difícil perquè com més complex és un sistema, menys probable és que aparega i persistesca»
Des d’aquesta perspectiva caldria centrar-se, llavors, a investigar distintes combinacions de components moleculars i processos d’interacció i transformació, fins trobar-ne algunes que (essent prebiòticament plausibles) assoliren una major cohesió, un manteniment dinàmic més estable, com a condició de possibilitat per a transicions ulteriors. Considerem, de fet, molt raonable plantejar que en certs entorns de la superfície terrestre fa uns 3.500 milions d’anys es degueren formar, impulsats per fonts d’energia externa (com el sol o l’energia geotèrmica), múltiples conjunts de reaccions químiques allunyades de l’equilibri en les quals es produïen gran diversitat de compostos, més o menys estables (figura 2). En aquest context, també és molt raonable assumir que part d’aquests compostos o agregats de compostos, si s’acumulen prou (sobre una superfície absorbent o en un microcompartiment), tindran un efecte sobre compostos i processos adjacents, de tal manera que potencialment contribuesquen al manteniment de tot el conjunt de reaccions (de la mateixa manera que, en un cicle autocatalític senzill, cada component del cicle contribueix a la síntesi de la resta). La idea és que aquestes formes de cohesió i automanteniment robust s’aconseguirien quan les influències entre els distints components en interacció tingueren un efecte de reforç mutu, evitant tendències naturals com la dispersió espacial o el decaïment d’estructures materials de no equilibri. No és senzill determinar el conjunt mínim de components o processos necessari per a obtenir una integració funcional autònoma, i per això és una qüestió que s’ha d’investigar empíricament. De totes maneres, la presència de mecanismes de control cinètic (catalitzadors) i espacial (compartiments), així com una coordinació adequada, sembla un requisit indispensable.
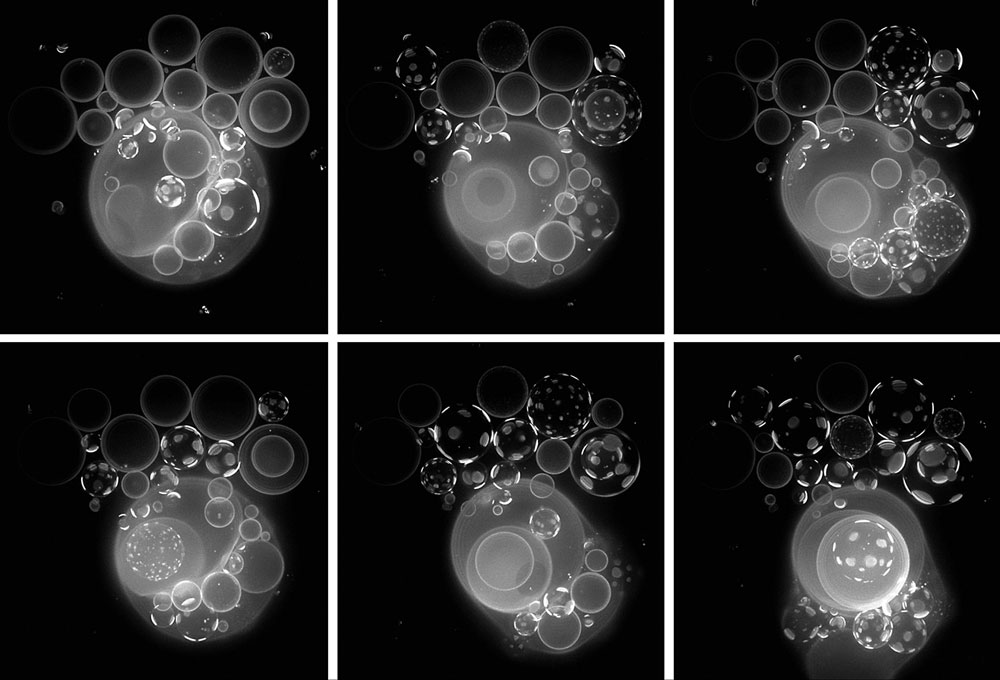
En tot cas, la qüestió més interessant és que aquests sistemes no sols serien viables en virtut d’unes determinades condicions externes (les quals sempre podrien fer-los desaparèixer, si foren prou adverses). Ells mateixos crearien i mantindrien gran part de les condicions que de fet possibiliten que es constituesquen i que progressen dinàmicament, ja que és la presència dels diversos components i la seua implicació en el conjunt de processos d’interacció i transformació el que fa possible la realització de la resta. Som ara en un escenari on opera un règim causal radicalment nou: grups de molècules generen un conjunt de patrons i estructures materials (altres molècules o agregats de molècules, com cadenes peptídiques amb capacitat catalítica o compartiments lipídics de permeabilitat selectiva –vegeu figura 3)– que constrenyen els processos i transformacions subjacents de manera que regeneren recursivament aquestes estructures i, a partir d’aquí, el conjunt de les interaccions que mantenen dinàmicament tot el sistema. Aquesta és una idea que altres autors (Kauffman i Clayton, 2006) han plasmat en termes de «cicles de constricció-treball», suggerint que les constriccions apareixen i es propaguen sempre que alguna configuració material en una part de l’univers estableix bucles no lineals i recursius d’aquest tipus, i que és un fenomen clau per a entendre l’origen de la vida. Es tracta, doncs, d’armar un entramat causal que, lluny de restringir o fitar l’espai de possibles estats dinàmics del sistema, permeta l’existència de nous estats estacionaris estables, i que reflectesca la major cohesió aconseguida entre els seus components.
Implicacions de l’enfocament alternatiu
Arribat aquest punt, estem ja en condicions de respondre al repte plantejat per Nagel. La matèria ha pogut transitar de l’univers fisicoquímic al biològic perquè ha trobat, com a resultat potser també d’esdeveniments contingents, formes estables d’organització dinàmica que sorgeixen de la seua capacitat d’«autoconstricció». Aquestes organitzacions, basades en un conjunt de bucles d’interacció recursiva, no lineal, no són exclusives de la biologia: en el món dels fenòmens físics han aparegut i apareixen espontàniament, com veiem en el cas dels huracans. Però en alguna etapa del procés prebiòtic, ja en un escenari d’interaccions i transformacions prou ric (és a dir, en un marc necessàriament químic), la innovació va ser que alguns sistemes mantinguts d’aquest tipus es van poder convertir en sistemes autoproductius, amb un major grau de cohesió interna i robustesa dinàmica, que van aconseguir generar múltiples constriccions mútuament dependents i aquestes, al seu torn, d’altres que es van fer cada vegada més complexes i diverses. La base d’aquesta complexitat creixent és que les constriccions generades dins del sistema obren de facto noves possibilitats organitzatives, ja que modulen selectivament les dinàmiques microscòpiques que tenen lloc en el si del sistema.
«Tot sistema biològic construeix una organització dinàmica a través de processos de síntesi i transformació de components, incloent-hi aquells que procuren les seues pròpies condicions de viabilitat i manteniment en el temps»
En aquest context, l’automanteniment pot començar a desplegar a més una nova dimensió: l’autoreproducció. En realitat, l’autoreproducció no és sinó una forma especial d’autoproducció. Però l’aparició de la vida requereix que els sistemes mantinguts adopten aquesta forma d’autoproducció que genera noves unitats, ja que només generant prou proliferació espacial i una continuïtat temporal indefinida (és a dir, una història oberta) basada en l’encadenament causal entre organitzacions individuals efímeres es pot assegurar la fixació i l’eventual increment de les innovacions que van anar apareixent per atzar. El que reté aquestes innovacions és simplement el fet que contribueixen al manteniment de les organitzacions individuals en què sorgeixen. Per tant, la seua funcionalitat i potencial avantatge selectiu mai podran explicar-se plenament fora del marc d’aquesta organització. I és això, juntament amb el fet que hi haja proliferació i transmissió hereditària, el que propicia el començament d’una forma primitiva d’evolució per selecció natural.
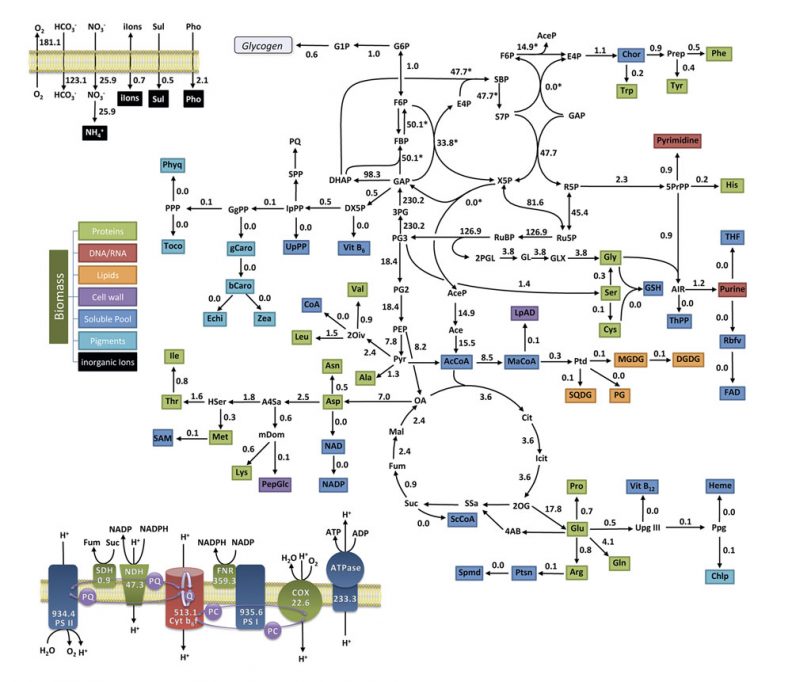
En resum, no hi ha «màgia» –com pretén Nagel– en l’origen de la vida. No hi ha màgia, però sí que som davant d’un fenomen extraordinàriament intricat i amb profundes implicacions, tant de científiques com de filosòfiques, ja que resulta, en un sentit important, inexplicable des de la lògica «habitual» que els físics i químics usen per a entendre els fenòmens no vivents. L’emergència de sistemes materials capaços d’automantenir-se de manera robusta i de dur a terme cicles reproductius estables (és a dir: metabolismes genèticament instruïts) comporta l’establiment de complexos bucles de transformació química (vegeu figura 4) que permeten la integració de mecanismes de regulació (basats en relacions de control jeràrquic entre diversos mòduls o subsistemes funcionals) junt amb mecanismes d’herència fiable (basats en relacions informacionals). Una teoria general i consistent sobre l’organització biològica, en el cas que siga factible elaborar-ne una, hauria d’explicar com poden sorgir sistemes naturals que generen i mantinguen aquest tipus d’arquitectures de control regulatiu i informacional, tenint en compte per fer-ho nous resultats provinents de la química/biologia de sistemes i la biologia sintètica.
«No hi ha màgia en l’origen de la vida, però sí que som davant d’un fenomen extraordinàriament intricat i amb profundes implicacions, tant de científiques com de filosòfiques»
Ací, sense pretendre avançar-nos a aquests resultats, hem tractat d’identificar alguna de les problemàtiques conceptuals fonamentals que intuïm que sorgiran en aquest interès i el tipus de plantejament teòric que es requerirà per a superar-les. La investigació sobre l’origen de vida, en la mesura que tracta de connectar el món de la física i la química amb el de la biologia, i en la mesura que està obligada a comprendre l’emergència i interrelació d’ambdues dimensions del fenomen (tant la seua organització individual –és a dir, l’aparició de metabolismes cel·lulars– com les seues capacitats evolutives a nivell col·lectiu –el procés històric de diversificació i increment de complexitat), constitueix un camp ineludible per a fer possible, si és pot, aquesta teoria en un futur. I en termes més amplis, per descomptat, també per a comprendre les relacions entre les distintes branques del coneixement científic ja establert. És a dir, fins que no resolguem la qüestió de l’origen no serem capaços de caracteritzar amb precisió la independència (o les formes d’interdependència) de la biologia en relació amb la física i la química.
AGRAÏMENTS
Els autors agraeixen la lectura del manuscrit i suggeriments de millora realitzats per dos revisors anònims. També volen deixar constància del suport a la seua labor d’investigació del Govern Basc (IT 590-13) i del Ministeri d’Economia i Competitivitat (FFI2011-25665 – KRM i AM), així com del d’Indústria i Innovació (BFU2012-39816-C02-02 – AM). A més, KRM és beneficiari d’ajudes en el marc de les Accions Europees COST CM1304 i TD1308.
REFERÈNCIES
Eigen, M., & Schuster, P. (1979). The Hypercycle: A principle of natural self-organization. Nova York: Springer.
Fry, I. (2000). The emergence of life on Earth: A historical and scientific overview. New Brunswick: Rutgers University Press.
Karsenti, E. (2008). Self-organization in cell biology. A brief history. Nature Reviews Molecular Cell Biology, 9, 255–262. doi: 10.1038/nrm2357
Kauffman, S., & Clayton, P. (2006). Agency, Emergence and Organization. Biology and Philosophy, 21, 501–521. doi: 10.1007/s10539-005-9003-9
Keller, E. F. (2007). The disappearance of function from ‘self-organizing systems’. En F. Boogerd, J. H. Bruggeman, H. V. Hofmeyr, & H. Westerhoff (Eds.), Systems Biology. Philosophical Foundations. (pp. 303–318). Dordrecht: Elsevier.
Lewontin, R. (1970). The Units of Selection. Annual Reviews of Ecology and Systematics, 1, 1–18. doi: 10.1146/annurev.es.01.110170.000245
Monod, J. (1970). Le hasard et la nécessité. Essai sur la philosophie naturelle de la biologie moderne. París: Éditions du Seuil.
Moreno, A. & Ruiz-Mirazo, K., 2009. The problem of the emergence of functional diversity in prebiotic evolution. Biology and Philosophy, 24(5), 585–605. doi: 10.1007/s10539-009-9178-6
Nagel, T. (2012). Mind and Cosmos. Why the Materialist Neo-Darwinian Conception of Nature is Almost Certainly False. Oxford: Oxford University Press,
Orgel, L. E. (1992). Molecular replication. Nature, 358, 203–209. doi: 10.1038/358203a0
Pattee, H. H. (1969). How does a molecule become a message? Developmental Biology Supplement, 3, 1–16.
Peretó, J., Bada, J. L., & Lazcano, A. (2009). Charles Darwin and the origin of life. Origins of Life and Evolution of Biospheres, 39(5), 395–406. doi: 10.1007/s11084-009-9172-7
Ruiz-Mirazo, K., & Moreno, A. (2012). Autonomy in evolution: From minimal to complex life. Synthese, 185(1): 21–52.
Simon, H. (1962). The architecture of complexity. Proceedings of the American Philosophical Society, 106, 467–482.
Szostak, J. W., 2012. The eightfold path to non-enzymatic RNA replication. Journal of Systems Chemistry, 3, 2. doi: 10.1186/1759-2208-3-2
Wicken, J. S., 1987. Evolution, thermodynamics and information. Extending the Darwinian program. Oxford: Oxford University Press.




