Invisible nets. The art of clarifying how many marine organisms there are, what they are and what they do. The greater part of the biomass of living creatures in the ocean is made up of microorganisms. Fundamental to explaining the way those ecosystems work they are responsible for most of the breathing taking place in marine waters and for the heterotrophic use of organic composts. All in all it was not until the end of the 20th century with the development of microscopic instrumentation that their task has been appreciated, putting into doubt the dominant belief that algae are the main generators of primary production (binding carbon). Nowadays, genomics has enabled us to gain insight into the functions of microorganisms that we did not know were present in the plankton.
No resulta gens evident quan ens mirem el mar. Però les aigües en les quals ens acabem de banyar són curulles de vida, i això que no s’hi veu cap peix. I és que la major part de la vida al mar és formada per organismes microscòpics visibles només amb aparellatge especialitzat. Si volguéssim reproduir a casa un mar artificial, primer caldria fer l’aigua salada: afegint uns 35 grams de sals per litre d’aigua. I després, afegir-hi els organismes. Quants? Doncs en un litre, 1.000 ciliats, 10.000 algues grans, un milió d’algues petites, mil milions de bacteris i deu mil milions de virus (a la fig. 1 n’hi ha alguns exemples). Déu-n’hi do la “gentada” que es troba en un litre d’aigua de mar. Dit d’una altra manera, hi ha tants organismes microscòpics en un litre d’aigua que si uns pescadors de teranyina volguessin reunir la mateixa quantitat d’anxoves que de bacteris hi ha en una gota, haurien de treballar ininterrompudament durant gairebé quatre anys. El nombre de microorganismes al mar és només comparable al nombre de grans de sorra que hi ha en una platja. En un litre d’aigua hi ha aproximadament tants microorganismes com grans en un metre cúbic de sorra. És a dir, molts. I malgrat ser tants, no els trobareu específicament anomenats a cap peixera de l’Aquàrium de Barcelona o l’Oceanogràfic de València.
I potser el més important és que aproximadament la meitat de la producció primària (la fixació de carboni) de la Terra és deguda a microorganismes. La major part de la respiració a les aigües marines és també deguda als microorganismes, i això és així perquè més del 90% de la biomassa d’éssers vius a l’oceà són microorganismes. L’alè de la Terra, allò que fa que diguem que la Terra és un planeta viu, és producte sobretot de l’activitat dels microorganismes marins.
Però la majoria dels microbis marins no creixen al laboratori, i per tant, ben poca cosa sabem de la diversitat que amaguen en una xarxa que ens resulta invisible. Només amb eines microscòpiques potents, tècniques de laboratori sensibles, i aparells moleculars que permetin estudiar el material genètic present a la mostra podrem conèixer qui són i què fan els habitants més abundants del medi marí. És interessant veure com ha anat evolucionant el nostre coneixement sobre els microorganismes marins, particularment els bacteris. El coneixement ha anat avançant a salts, impulsats sovint per l’accessibilitat dels oceanògrafs marins a tècniques noves i, en fer-ho, sovint ha generat sorpreses, han aparegut coses inesperades que han fet trontollar i refer els paradigmes que fins al moment dominaven. D’això en parlarem aquí.
Fins fa tres dècades, la imatge que es tenia de les xarxes tròfiques marines era la representada a la fig. 2a: la producció primària, la generaven les algues de mesures situades entre 2 i 200 µm (mil·lèsimes de mm), i aquesta producció primària representava l’aliment dels protistes ciliats (mides: 30 – 500 µm) i dels copèpodes (el zooplàncton). Alhora, els peixos s’alimentaven d’aquests copèpodes. En aquest esquema, els microorganismes eren totalment innecessaris per a explicar el funcionament de l’oceà. En part això era causat perquè les tècniques que s’usaven en aquell moment per determinar l’abundància de bacteris provenien de la microbiologia mèdica: medis de cultiu en placa molt rics en components orgànics. Però diversos investigadors havien vist que alguna cosa no lligava. Així com una bona part de la producció primària quedava retinguda per filtres de 20 µm (és a dir, era portada a terme per organismes més grans d’aquesta mesura), la major part de la respiració i de la utilització heterotròfica de compostos orgànics tenia lloc en organismes menors d’1 µm. És a dir, havia de ser deguda als bacteris. Cap al 1975 (vegeu taula 1) es van introduir a la biologia marina tècniques de tinció de DNA i d’epifluorescència, i immediatament mostràrem que al mar hi havia si fa no fa un milió de bacteris per mil·lilitre.
| Data aprox. | Descobriment o canvi conceptual proposat |
| 1977 | Hi ha molt més bacteris al mar del que pensàvem. |
| 1980 | Aquests bacteris creixen i es reprodueixen activament. |
| 1982 | Els bacteris són depredats activament, sobretot per flagel·lats heterotròfics. |
| 1986-1988 | Flagel·lats i ciliats poden ser mixotròfics. |
| 1988 | Descobriment de Prochlorococcus (alguns bacteris fan la funció d’algues). |
| 1990 | Els virus representen un paper important a les xarxes tròfiques planctòniques. |
| 1990 | La major part de bacteris marins són totalment desconeguts. |
| 1992-1994 | Hi ha arqueus al plàncton marí, fins i tot a l’Antàrtida. |
| 1995 | No tots els bacteris que veiem són vius ni estan actius. |
| 1997 | La respiració dels bacteris marins supera i la producció primària en mars oligotròfics. |
| 1998-2000 | Desenvolupament de tècniques per a l’anàlisi cel·lular a escala individual. |
| 2000 | Descobriment d’organismes fotoheterotròfics al mar. |
| 2001 | Els arqueus dominen al mar profund aeròbic i relativament temperat. |
| 2001 | Nous cianobacteris unicel·lulars que també fixen nitrogen. |
| 2001 | Es descobreix la diversitat amagada en els petits eucarionts marins. |
| 2002 | S’aconsegueix aïllar un SAR11 en cultiu pur. |
| 2003 | Se seqüencien els genomes de Prochlorococcus y Synechococcus. |
| 2003 | S’aïllen virus de Prochlorococcus i de diatomes. |
| 2004 | Reconstrucció del metagenoma del mar dels Sargassos. |
| 2005 | Els arqueus marins tenen metabolismes quimioheterotròfics. |
Taula 1. Algunes fites rellevants en el coneixement de l’estructura i el funcionament de les xarxes tròfiques microbianes marines.
Malgrat l’evidència directa, però, no va ser fins uns quants anys després, a començament dels vuitanta, que, amb l’ajut de traçadors radioactius, es va demostrar que els bacteris no sols eren abundants, sinó que també eren una part dinàmica de l’ecosistema marí: creixien i es duplicaven ràpidament i la seva producció podia arribar al 30% de la producció primària. Els bacteris es desenvolupaven usant la matèria orgànica dissolta i particulada produïda pels productors primaris, les algues, però també utilitzant matèria orgànica d’origen terrestre, i degradant fragments orgànics provinents de les restes de l’alimentació de copèpodes sobre algues i peixos sobre copèpodes. Quan es va començar a veure que els bacteris marins eren molt abundants, creixien, es reproduïen i participaven activament en la producció de l’ecosistema i en els cicles biogeoquímics de l’oceà, va ser quan la major part d’oceanògrafs van començar a pensar que aquella xarxa tròfica de la fig. 2a potser no era la dominant a l’oceà.


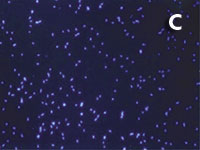
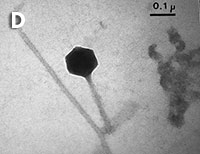
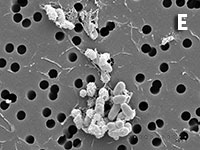
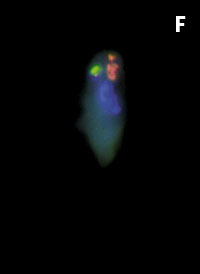
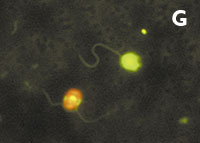
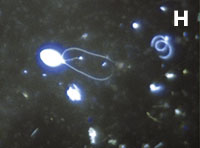
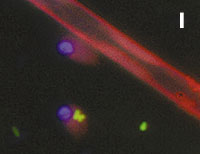
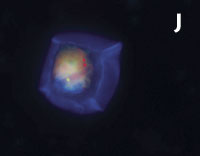
Figura 1. Imatges del mostreig de comunitats planctòniques marines al Pacífic (A) i a l’Antàrtida (B). L’aparell és una roseta d’ampolles oceanogràfiques agafades a un sensor CTD (de conductivitat, temperatura i densitat). Imatges de diversos microorganismes observats amb diferents tipus de microscòpia. C) Bacteris tenyits amb DAPI i observats amb epifluorescència de radiació ultraviolada. D) Imatge d’un bacteriòfag amb microscòpia electrònica de transmissió. E) Bacteris observats amb microscòpia electrònica d’escandallatge. Els porus del filtre fan 0,2 µm de diàmetre. F) Un ciliat, amb el nucli tenyit de blau, que ha menjat bacteris (grocs) i Synechococcus (taronja). G) Un flagel·lat heterotròfic (groc) i un de fototròfic (vermellós) en mostres de l’Àrtic. H) Un flagel·lat heterotròfic. I) Flagel·lats fototròfics (vermells de la clorofil·la i blau de la tinció del nucli) un dels quals ha ingerit bacteris (groc) i és per tant mixotròfic. També s’hi veu una diatomea. J) Un dinoflagel·lat amb el nucli tenyit de blau i vermell de la clorofil·la. (Imatges F, I i J, de Fernando Unrein; C, de Klaus Jürgens; G, de Dolors Vaqué; D, de Núria Guixa-Boixereu; E, de José M. Fortuño.)
Si els bacteris creixien, encara que quan es prenien mostres al llarg del temps no es veia que n’augmentés la concentració, era evident que calia un mecanisme que “eliminés” bacteris del medi, i aquest mecanisme es va trobar a començament dels anys vuitanta, quan es va fer evident que, a part de bacteris, a l’oceà hi havia també molts protists unicel·lulars, flagel·lats heterotròfics en concentracions properes als 1.000 organismes per mil·lilitre que depredaven els bacteris. Alhora, aquests flagel·lats servien d’aliment al zooplàncton, de manera que “enllaçaven” el que es va anomenar la xarxa tròfica microbiana (bacteris que utilitzen matèria orgànica i que són aliment de protists unicel·lulars) amb la xarxa tròfica clàssica (el zooplàncton que és aliment de peixos). D’aquí que es parlés del bucle microbià (microbial loop) pel qual el carboni que hom creia que es perdia del sistema, de fet era reintroduït per acció dels bacteris i els flagel·lats heterotròfics (fig. 2b).
Quan ja es pensava que es coneixia l’estructura de les xarxes tròfiques marines i els principals fluxos biològics entre components de la xarxa, tres descobriments de finals dels anys vuitanta i començament del noranta van fer adonar els biòlegs marins que la “xarxa invisible” de microorganismes al mar encara amagava secrets rellevants. Primer, es va veure que una bona part de protists marins presumptament heterotròfics de fet eren mixotròfics. És a dir, tant podien fer la fotosíntesi –i per tant tenien pigments i ecològicament se’ls podia considerar com a algues– com podien comportar-se com a depredadors de bacteris i, per tant, eren alhora autotròfics i heterotròfics. Una estratègia que té sentit si es pensa que el mar és un ambient relativament pobre en nutrients i que ser capaços d’usar totes les possibilitats metabòliques permet més possibilitats per a sobreviure. Pràcticament alhora, i gràcies a la introducció de la cromatografia líquida d’alta pressió en l’estudi dels pigments de les algues, i a la de la citometria de flux –tecnologia originària de la biomedicina clínica–, es va descobrir que al mar hi havia un organisme, de mesura i estructura cel·lular bacteriana (0,8 µm de diàmetre), que era fotosintètic i extremadament abundant. El Prochlorococcus, que així és com s’anomena, és tan abundant i tan estès pels mars temperats i càlids de l’oceà que hi ha qui ha dit que probablement és l’organisme fotosintètic més abundant de la Terra. Us imagineu que ara, de sobte, algú descobrís als boscos tropicals una espècie de planta que resultés ser la més abundosa i estigués estesa arreu de la Terra? Sorprendria, oi? Doncs així ens vam quedar quan a finals dels vuitanta es va conèixer aquest organisme. Finalment, a començament dels noranta es va descobrir que els microorganismes marins, bacteris i protists, no se salvaven de ser atacats per virus i la mortalitat causada pels virus podia explicar moments en els quals hi havia alta producció bacteriana però poca mortalitat per protists. Els virus també atacaven els bacteris i podien ser responsables de fins a la meitat de la mortalitat bacteriana. Malgrat això, fins fa dos anys encara no es coneixia cap virus que ataqués Prochlorococcus, i fins fa molt poc no se’n coneixien que ataquessin les diatomees, un grup d’algues eucarionts que dominen les xarxes tròfiques de les regions oceàniques amb més nutrients, més producció primària i més producció pesquera.
La presència d’organismes amb la funció ecològica de les algues, però de mida i estructura bacteriana, i el paper dels virus fan replantejar la xarxa tròfica microbiana (fig. 2b), ampliant els enllaços d’aquesta malla invisible (fig. 2c). La dècada dels noranta va portar també altres sacsejades al paradigma, com quan es va fer evident que no tots els bacteris del plàncton eren vius, sinó que una fracció dels que havíem comptat des de sempre eren inactius, o fins i tot alguns eren morts (o ni tan sols eren bacteris, sinó restes de membranes de bacteris lisats per virus o mig menjats per flagel·lats). Un altre treball rellevant, publicat el 1997, va donar a conèixer que l’activitat respiratòria dels bacteris podia ser, en determinats ambients marins, més elevada que l’activitat dels productors primaris, de manera que el balanç net metabòlic de l’ecosistema era de producció de CO2, i no de consum d’aquest CO2 i producció d’oxigen com es pensava fins al moment. Aquest estudi, i d’altres que el van seguir, van encetar una polèmica que encara dura, al voltant de si el balanç net metabòlic de l’oceà és heterotròfic o autotròfic.
Però un dels avenços més significatius va tenir lloc quan, l’any 1990, tècniques manllevades dels laboratoris de biologia molecular es van aplicar per primera vegada a l’estudi de la diversitat microbiana marina. El resultat dels primers estudis va tornar a trencar paradigmes: la major part dels bacteris que es trobaven al mar no havien estat mai aïllats en cultiu (i, alhora, la majoria dels que s’havien aïllat del mar, eren irrellevants a la natura). No sols això, sinó que alguns grups concrets d’organismes, com els anomenats SAR11, un subgrup dels alfa-proteobacteris, apareixien a la major part d’oceans que s’estudiaven. De fet, encara ara hi ha molts grups d’organismes marins que no s’han aconseguit aïllar i que no se sap quina mena de metabolisme tenen, ni quina és la seva funció a l’oceà (fig. 4). Una altra sorpresa va arribar tot seguit. Els arqueobacteris són organismes procarionts aparentment semblants als bacteris, però que perta nyen a una línia evolutiva diferent, de fet entremig dels bacteris i dels eucarionts. Tots els arqueobacteris que es coneixien fins al moment havien estat aïllats de llocs hipersalins, d’ambients de temperatura molt elevada, o eren simbionts d’altres organismes. Per tant, no s’esperava que n’hi hagués al mar, i encara menys que n’hi hagués als mars antàrtics amb molt d’oxigen i temperatures fregant els zero graus. I vet aquí que l’any 1994 es va trobar que els arqueus podien representar el 20% dels procarionts antàrtics. I l’any 2001 es va saber que els arqueus dominaven el mar profund, per sota dels 2.000 metres, ambients freds i plens d’oxigen. I encara més, gràcies al desenvolupament de tècniques que permeten “interrogar” cèl·lules individuals de microorganismes (com ara la microautoradiografia, usant sondes radioactives combinades amb sondes filogenètiques), fa poc hem sabut que, molt possiblement, els arqueus marins tenen metabolismes quimioheterotròfics, que fixen CO2 amb energia obtinguda de reaccions químiques d’oxidació-reducció. L’aplicació de les mateixes tècniques de biologia molecular que van permetre “obrir” la caixa negra dels bacteris (a partir de l’any 1990) i dels arqueus (a partir del 1992) per saber de quins elements estava composta es va fer l’any 2001 als protists marins. També aquí vam veure que hi havia grups que no es coneixien abans i que semblaven dominar en molts oceans diferents. I ens adonàrem que sabíem molt poc del metabolisme d’aquests organismes.
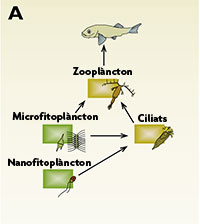
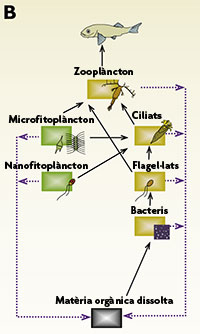
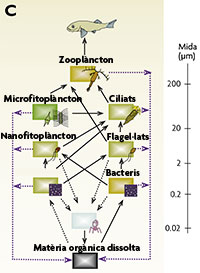
Figura 2. Tres models de xarxa tròfica planctònica amb indicació dels fluxos de carboni particulat (ratlles contínues) i de carboni orgànic dissolt i nutrients inorgànics (ratlles blaves discontínues). El model de l’esquerra (A) és el vigent abans de 1970, el central (B) exemplifica el concepte de “bucle microbià”, i el de la dreta (C) és el model que es creu correcte actualment. Les escales de la dreta mostren la mesura mitjana aproximada dels organismes, i els colors de les capses reflecteixen si es tracta de grups autotròfics (verd), heterotròfics (marró vermellós) o mixotròfics (marró verdós).
Els darrers anys han portat nous descobriments que, sovint, ens han fet canviar algunes idees que consideràvem fixes. Per exemple, hom creia que era impossible que hi hagués cianobacteris unicel·lulars que fessin fotosíntesi i fixació de nitrogen, perquè els dos metabolismes es creia que eren incompatibles en la mateixa cèl·lula –l’oxigen produït a la fotosíntesi inhibeix els enzims de la fixació de nitrogen– i els cianobacteris filamentosos segreguen les dues funcions en cèl·lules especialitzades diferents. Doncs bé, l’any 2001 es van trobar cianobacteris unicel·lulars i fixadors de nitrogen. A les tècniques de biologia molecular s’hi han afegit avenços en les tècniques de cultiu i en les d’aïllament i anàlisi de cèl·lules individuals. Així, l’any 2002 es presentava, per primera vegada, l’aïllament en cultiu pur d’un representant dels SAR11, anomenat Pelagibacter ubique, un dels bacteris marins dels quals es trobava sempre el DNA, però que no se’n sabia res de la seva morfologia, ni del metabolisme, ni de l’ecologia. Resulta ser un organisme típicament oligotrof: és petit, creix amb poca concentració de nutrients (una alta concentració els inhibeix), i no arriba mai a concentracions elevades en cultiu.
El darrer avenç rellevant en l’estudi dels microorganismes marins ha estat obtingut per l’aplicació de tècniques de seqüenciació ràpida de tot el genoma dels organismes que estan en cultiu, o de tot el contingut genètic present en una mostra de plàncton. Tot i que la seqüenciació era possible des de feia temps, no es va emprar per a diversos organismes sencers fins als avenços portats pel projecte de seqüenciació del genoma humà. Actualment s’estan seqüenciant uns quants centenars de microorganismes, inclosos alguns dels que es creu que són importants al medi marí. Conèixer el genoma dels microorganismes ens permet, d’una banda, entendre l’eco logia dels organismes: per què es troben on es troben, per què són dominants en alguns ambients i no en d’altres (requadre pàg. 78). Però, de l’altra, la genòmica ens obre la porta al coneixement de funcions dels microorganismes que ignoràvem que fossin presents al plàncton. Per tal de fer això se sol recollir molta biomassa de bacteris (uns 1.000 litres, és a dir, uns 1.000 bilions de microorganismes), es pren tot el DNA de la comunitat i se seqüencia. Si es té sort, es pot trobar gens que codifiquen per a funcions desconegudes al plàncton en el mateix fragment que un gen del RNA16S, que es pot seqüenciar i utilitzar com a marcador d’identitat filogenètica. L’anàlisi detallada que Oded Béjà i els seus col·laboradors van fer d’un metagenoma marí el va fer trobar gens de proteorodopsines (unes proteïnes que capten la llum del Sol i que alguns bacteris usen per generar ATP, energia, a partir de la llum) en organismes d’un grup que se sabia que existia al plàncton, però que no se sabia què hi feia (els SAR86). Ni la funció ecològica (utilització de la llum per a generar ATP) ni la seva adscripció a aquest grup concret d’organismes era previsible. Els seus estudis, juntament amb altres descobertes recents, han portat a un canvi de paradigma del funcionament de l’ecosistema planctònic (fig. 3). Abans es creia que només hi havia dues formes de convertir carboni dissolt en particulat: la fotoautotròfia practicada per algues i bacteris que utilitzen la llum del Sol per fixar carboni inorgànic, i la quimioheterotròfia, és a dir, la capacitat dels bacteris per usar matèria orgànica dissolta (provinent del mateix plàncton però també d’altres ecosistemes). La resta d’organismes depreden sobre els qui fan aquests dos models d’incorporació d’energia a les xarxes tròfiques. Doncs bé, ara caldria incorporar-hi una tercera via: organismes que usen matèria orgànica dissolta però alhora completen les seves necessitats energètiques gràcies a pigments i proteïnes (la proteorodopsina), que capten l’energia del Sol i l’usen per generar ATP, la moneda energètica dels éssers vius. L’estratègia és semblant a la dels protists mixotrofs, adequada per a la vida en un medi diluït com és la capa fòtica dels oceans.
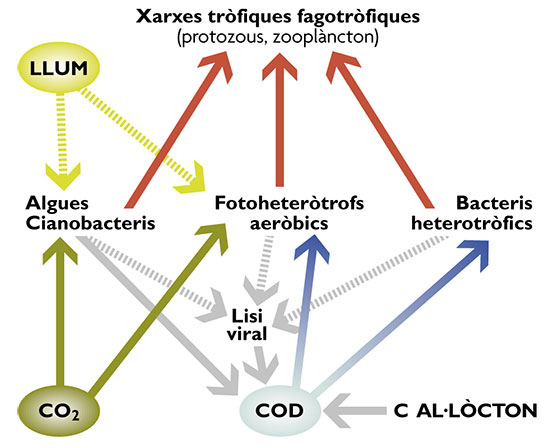
Figura 3. Els metabolismes bacterians a l’oceà il·luminat: hom creia que només hi havia fotoautòtrofs, que incorporen CO2 amb la llum, i quimioheteròtrofs, que utilitzen carboni orgànic dissolt (COD). Hi ha una tercera via, que usa alhora llum per generar energia, però que obté el carboni del reservori de carboni dissolt. (Modificat de Fenchel, T. 2001 Science 292: 2444-2445.)
Entitats com la Gordon and Betty Moore Foundation americana (liderada per un dels fundadors d’Intel), han decidit invertir en l’estudi dels genomes dels microorganismes marins, convençuts que obtindran grans descobriments. Les comunitats diverses de bacteris, arqueus i protists, no són sinó fàbriques on tenen lloc les reaccions químiques que determinen els cicles biogeoquímics de la Terra. També Craig C. Venter, l’inusual científic que competí des de l’empresa privada amb el consorci públic per a la seqüenciació del genoma humà, ha decidit “seqüenciar l’oceà”. Si ell hi olora grans oportunitats per a nous descobriments, alguns d’ells potencialment rendibles econòmicament, alguna cosa deu haver-hi. El fet és que podem preveure que aquest article serà vell d’aquí uns pocs anys, car sabrem moltes més coses sobre el funcionament de l’oceà que ara no podem ni intuir. Són coses que estan amagades a la xarxa invisible dels microorganismes. Les sorpreses en els paradigmes de l’estructura i el funcionament de l’ecosistema planctònic continuaran i caldrà estar-hi alerta.
Bibliografia Complementària
Institut de Ciències del Mar de Barcelona: Web de divulgació de l’Insti tut de Ciències del Mar de Barcelona.
Eosll Publishers: Enciclopèdia en xarxa sobre temes d’ecologia. És en anglès i cal registrar-s’hi.
J. Craig Venter Institute (2005): L’expedició de C. Venter per a l’estudi genòmic de l’oceà.
González, J., (en premsa) 2005. El bacteriopláncton en los océanos. Investigación y Ciencia.
Massana, R., 2005. “El picoplàncton marí, reserva d’organismes i funcions”, Omnis Cellula (en premsa).