Does the language faculty lie in our genes? Traditionally, language was considered as one of the most representative examples of cultural transmission (as opposed to biological transmission), a purely cultural phenomenon. From the second half of the twentieth century, though, that conception was dramatically questioned, by asserting that the language faculty is indeed biologically rooted. The objective of this article is threefold: first, it provides a brief picture of the arguments, mainly arising from linguistic development, which point to such a biological character. Further, it is argued that the difference between genotype and phenotype also applies to the linguistic field. Finally, evidence which crucially shows the genetic roots of language is reviewed and discussed: the recent discovery of an autosomal dominant gene (named “FOXP2”), whose disruption triggers severe language disorders.
Durant llarg temps, es va establir una contundent contraposició entre naturalesa (nature) i criança (nurture); mentre que l’ésser humà era el (únic) representant del segon domini, les altres espècies quedaven confinades en l’àmbit natural. Segons aquesta visió, la nostra espècie, a diferència de la resta, havia transcendit els factors genètics; així, la primacia dels gens s’equiparava amb un veto de qualsevol tipus d’aprenentatge, mentre que alliberar-se d’aquesta primacia atorgava una capacitat il·limitada d’aprenentatge; precisament, la imatge tradicional de l’ésser humà (fortament antropocèntrica) era el millor exemple del que es pot obtenir quan una espècie s’allibera del control dels instints. Instint, doncs, s’oposava a aprenentatge, i hi ha una bretxa insalvable entre els dos plans.
L’exemple paradigmàtic de la primacia de la criança en la nostra espècie va ser el llenguatge. Aquesta visió, que va trobar especial ressò en la tradició estructural-antropològica, és caracteritzada per Baker (1996: 507): “Aquesta tradició considera el llenguatge com un fenomen cultural; per tant, les llengües difereixen per les mateixes raons per què difereixen les cultures. En concret, poden ser modelades en vies importants per la visió del món, els models d’interacció i la història particular d’un grup concret de persones. Com que aquests factors culturals poden ser molt diferents, en la mateixa mesura les llengües seran molt diferents”. En suma, el llenguatge s’erigia en el principal estendard dels progressos que l’ésser humà havia aconseguit mitjançant la cultura.
Aquesta concepció responia a una motivació global molt clara (encara persistent): la resistència per a acceptar una genètica del comportament; enfront del paper diàfan dels gens en el pla fisiològic, la idea que també són rellevants en la conducta és polèmica, perquè sol identificar-se amb el determinisme.
No obstant això, aquesta concepció va començar a qüestionar-se en la segona meitat del segle XX en nombrosos dominis, lingüística inclosa, de manera que es va anar rebutjant el caràcter enfrontat de naturalesa i criança, i es fa afavorir en lloc seu una integració dels efectes d’ambdues. El biòleg Michael Ruse (1986: 183) assenyala que “des d’una perspectiva darwinista general, suposar que una cultura està aïllada al cim dels gens és ineficient i perillós per a ambdues parts. La ment com a tabula rasa requereix un cervell amb una gran capacitat en blanc […]. Aquesta receptivitat total requeriria probablement una capacitat cranial molt major de la que ara posseïm. Aquest requisit converteix la hipòtesi en altament improbable”. Per tant, la possessió, adquirida evolutivament, de diferents capacitats d’aprenentatge augmenta la fitness (eficàcia biològica) dels organismes, ja que l’aprenentatge és un mitjà de què disposen els gens per a ajustar-se a les exigències canviants de l’entorn. Wilson (1978: 99-100) defensa clarament la complementarietat d’allò innat i d’allò après: “El potencial d’aprenentatge de cada espècie sembla estar totalment programat per l’estructura del seu cervell, la seqüència de les descàrregues de les seues hormones i, finalment, pels seus gens. Cada espècie animal està ‘preparada’ per a aprendre certs estímuls, impedida d’aprendre’n altres i neutral respecte a uns altres més”.
Les dues òptiques assenyalades, la tradicional i la moderna, responen a dues concepcions diferents sobre l’aprenentatge: aprenentatge per instrucció i per selecció. Segons la primera, corresponent a l’òptica tradicional, el mecanisme d’aprenentatge consisteix a traspassar informació des de l’exterior (entorn) a l’interior de l’organisme, de manera que aquest manca de tota capacitat prèvia i és construït de manera gradual per l’experiència, segons va interioritzant els estímuls pertinents. Enfront de l’òptica tradicional, l’aprenentatge per selecció implica que l’organisme desenvolupa trets compatibles amb les capacitats preexistents del programa genètic. La diferència entre ambdues nocions és acusada: mentre que l’aprenentatge per instrucció descansa únicament en el paper de l’experiència, encarregada de configurar per complet l’ésser, l’aprenentatge per selecció relativitza aquest paper; sense negar òbviament la seua importància, la funció de l’experiència és disparar o activar trets innats específics de domini que hi preexisteixen.
La substitució d’un paradigma per un altre permet inserir l’ésser humà en una posició realista, d’acord amb la resta d’espècies (v. a aquest respecte López García, 2002, que remet el procés comunicatiu al nivell cel·lular mateix): des de l’òptica tradicional, l’espècie humana era una mena d’illa en meitat del regne animal; illa merament cultural, mancada de qualsevol instint, ubicada, però, al si d’una naturalesa plena d’espècies amb instints específics. Com per art de màgia, ens hauríem alliberat del predomini dels instints i així vam accedir a un aprenentatge il·limitat.
També el llenguatge participa d’aquest canvi de perspectiva. La idea segons la qual el llenguatge implicava un fenomen cultural per excel·lència deixava sense explicar per què només el nostre sistema, enfront de la resta de la comunicació animal, estava lliure de factors genètics. De nou ens topem amb aquest caràcter d’illa al mig del mar de la naturalesa que ens vam atorgar durant llarg temps. Avui dia, però, hi ha nombroses proves sobre l’existència d’un aprenentatge “preparat” per al llenguatge; en suma, sobre l’existència de factors innats implicats en la seua adquisició. Els repasse breument (v. Lorenzo, Longa, 2003: cap. 2, per a una visió detallada).
Indicis de la predeterminació biològica del llenguatge
Els arguments esgrimits (especialment, per l’innatisme de Noam Chomsky) sobre aquesta predeterminació biològica, es deriven bàsicament de l’anàlisi de l’adquisició del llenguatge en el nen. Amb això, l’estudi del llenguatge està en paral·lel amb el d’altres dominis cognitius humans, també centrats en el desenvolupament com a font central d’informació. Aquests arguments dibuixen el següent panorama.
«Tots els grups i societats humanes posseeixen llenguatge i només la nostra espècie el posseeix. A més, totes les llengües s’han revelat com a sistemes d’idèntica complexitat, amb independència del grau de desenvolupament o de l’aïllament d’una societat»
Tots els grups i societats humanes posseeixen llenguatge i només la nostra espècie el posseeix. A més, totes les llengües, plasmacions de la facultat del llenguatge, s’han revelat com a sistemes d’idèntica complexitat, amb independència del grau de desenvolupament o de l’aïllament d’una societat. D’altra banda, hi ha correlats neurals estretament lligats a aquesta capacitat. Auditivament, estem preparats per a discriminar sons lingüístics amb molta major eficiència que els no lingüístics, en una proporció de tres a un. El llenguatge està sotmès a un període crític: tan sols es pot adquirir fins una determinada edat; més enllà de la qual, l’exposició a l’experiència només capacita per a adquirir un protollenguatge privat de les propietats centrals del llenguatge. Hi ha una gran diferència entre l’adquisició de la llengua nativa i la de segones llengües: mentre que tots arribem (patologies a banda) a dominar perfectament la nativa, independentment del nivell d’intel·ligència, educatiu, etc., l’adquisició d’una segona llengua, superat el període crític, ha de fer-se amb esforç i produeix gran variació en els resultats. El nen adquireix en edat molt primerenca un sistema de gran complexitat com el llenguatge (als tres anys ja en posseeix un gran domini) en el temps en què és incapaç d’afrontar tasques cognitives molt més simples. El llenguatge i la resta de la cognició són entitats dissociables: una deficiència cognitiva pot acompanyar una funció lingüística no afectada i viceversa.
Els nens travessen per etapes i ritmes que mostren gran concordança amb independència de la llengua adquirida (també en llengües de signes) i la “quantitat” de l’experiència no és rellevant: algunes cultures, com la nostra, tenen un estil de parla dedicat al nen (maternès), mentre que altres ignoren el nen fins que pot parlar amb solvència; a pesar d’això, en ambdós casos adquireixen el llenguatge sense diferències temporals. El nen pot desenvolupar sistemes gramaticals complexos a partir d’una evidència pobra, oral o gestual. Els enunciats lingüístics, rebuts linealment, oculten l’estructura formal de les oracions, que és jeràrquica; a pesar d’això, el xiquet l’aprèn molt ràpidament sense cap tipus d’instrucció i desenvolupa un sistema intricat i mancat de correspondència amb els senyals a què està sotmès (pobresa de l’estímul).
«Tots aquests fets, presos en conjunt, fan inviable sostenir que aprendre a parlar implica una mera interiorització d’hàbits sorgits en el marc cultural; ben al contrari, apunten que el llenguatge és un tret biològicament assentat»
Tots aquests fets, presos en conjunt, fan inviable sostenir que aprendre a parlar implica una mera interiorització d’hàbits sorgits en el marc cultural; ben al contrari, apunten que el llenguatge és un tret biològicament assentat.
Genotip i fenotip
Si el llenguatge és una facultat amb un suport biològic més que un tret cultural (encara que òbviament el factor cultural siga rellevant per a adquirir una llengua concreta), hem d’aplicar la lògica que subjau a l’aprenentatge per selecció. S’hi han de diferenciar dos components (els efectes dels quals se sumen): l’estat inicial i l’experiència, que activa el primer. La interacció d’ambdós factors produeix un estat estable donat (el tret en qüestió). Estenent aquesta visió al llenguatge, l’estat inicial és únic per a l’espècie, invariable entre els individus amb independència de la llengua que adquiresquen (òbviament, és inviable postular diferents assentaments genètics per a diferents llengües). Aquest estat inicial comú a l’espècie, sotmès a una experiència canviant (l’exposició a una o una altra llengua), determina l’estat estable (el coneixement del turc, del gallec o de l’hindi). La missió de l’estat inicial és configurar la forma d’una llengua humana possible, els trets abstractes de disseny de qualsevol plasmació del llenguatge, actuant així com a guia del procés d’adquisició, que motiva que el nen explore certes opcions però que en descarte automàticament d’altres sense considerar-les; d’aquí se deriva la rapidesa d’adquisició, al costat de la resta de característiques assenyalades.
El genotip, conformat pel material genètic, determina el rang potencial d’adaptacions funcionals a l’entorn que un organisme pot desenvolupar segons entre en contacte amb diferents tipus d’ambients. Per la seua banda, el fenotip són les propietats morfologicoestructurals i funcionals desenvolupades en funció de la interacció entre els gens de l’organisme i el seu entorn. Així, el genotip, programa intern de desenvolupament, es plasma en un fenotip específic segons determinacions ambientals específiques. És factible traslladar ambdues nocions al llenguatge: els trets assenyalats sobre l’adquisició apunten a un genotip o estat inicial comú, disparat per l’experiència; aquesta no omple un organismo “nu”, sinó que activa respostes internes a l’organisme. Prenent un paral·lelisme, la formació de l’eix de l’estructura de fucus (un tipus d’alga) necessita un estímul extern, com la llum, però aquest estímul no és el responsable de l’esmentada formació, que depèn de l’organització interna de l’ou. Per tant, la llum és un disparador que posa en marxa una cosa que està preparada per a eixir.
«En l’estat inicial lingüístic no s’especifiquen les propietats de les diferents llengües; el paper d’aquest estat és descartar per endavant la possibilitat que es desenvolupen propietats formals concebibles des de premisses lògiques, però incompatibles amb ell»
L’estat inicial comú a totes les llengües, la dotació genètica del llenguatge, no es troba de segur directament codificada en el genoma, encara que sí que determinada per la seua composició. Monod (1970: 100) va assenyalar que, per a alguns, era una contradicció afirmar que el genoma defineix completament la funció d’una proteïna i al mateix temps que el contingut informatiu d’aquesta funció siga molt més ric que el mateix contingut del genoma. No obstant això, l’enriquiment informatiu, des de la codificació genètica fins a la seua expressió final, implica realitzar una estructura somàtica particular entre moltes altres concebibles a priori, però irrealitzables a partir de les mencionades condicions inicials. Per això, la informació final no té per què estar especificada en el genoma, que no obstant això descarta altres opcions de desenvolupament. De manera semblant, en l’estat inicial lingüístic no s’especifiquen les propietats de les diferents llengües; el paper d’aquest estat és descartar per endavant la possibilitat que es desenvolupen propietats formals concebibles des de premisses lògiques, però incompatibles amb ell. Els diferents estats finals obtinguts o llengües són sistemes diferenciats entre si, però subjectes a un disseny o esquematisme comú, que constitueixen estats alternatius de desenvolupament.
Un panorama com aquest es desprèn de les dades sobre l’adquisició, que apunten a una clara base innata. En tot cas, no hem d’equiparar “innat” amb estrictament genètic: al costat hi ha els processos d’epigènesi, interacció entre gens i entorn, que suposen processos d’emergència no ben coneguts. De fet, és l’epigènesi, especialment rica en l’ésser humà, la que motiva l’absència d’una relació tancada entre gens i comportament, cosa que dóna un enorme marge de maniobra a l’organisme.
Les bases genètiques del llenguatge
Si la facultat del llenguatge té suport innat, s’ha de transmetre per herència. Com és ben sabut, els gens no poden codificar directament principis funcionals (aquesta és, de fet, la principal reticència que desperta la genètica del comportament); la seua missió és codificar la síntesi de les proteïnes. Jackendoff (2002: 90) assenyala que les proteïnes estableixen paràmetres de desenvolupament cerebral encara desconeguts; per això, per ara, no hi ha esperança de comprendre els mecanismes indirectes que operen en la transmissió genètica del llenguatge (tampoc la de la resta de nivells d’organització funcional)
|
Per què l’al·lel mutat és autosòmic i dominant?
Gopnik va afirmar que el gen implicat en el trastorn específic del llenguatge és autosòmic (lligat a un dels 22 parells de cromosomes no sexuals) i dominant. El trastorn afectava aproximadament el 50% dels membres d’una família KE (16 de 31). Aquest percentatge impossibilita que l’al·lel en qüestió siga de naturalesa autosòmica recessiva; en aquest hipotètic cas, la probabilitat de transmetre el gen patogen de cada progenitor és del 50%, i per això, la de rebre’l dels dos progenitors i manifestar els efectes seria del 25%, no del 50%, com succeeix en la família. Tampoc es pot considerar recessiu, però lligat al cromosoma X, perquè en aquest cas, el baró podria manifestar els seus efectes amb una probabilitat del 50% a condició que només la mare fóra portadora; no obstant això, la probabilitat en aquest cas per a la dona continua situada en el 25%. Enfront d’això, els casos de dones afectades en la família són superiors als d’homes. De les dues possibilitats restants, cal descartar la seua naturalesa dominant però lligada al cromosoma X, perquè aquest supòsit implicaria que no hauria d’haver-hi dones afectades només per transmissió paterna (el gen sa de la mare bastaria per a ocultar l’efecte); no obstant això, van ser descrits dos d’aquests casos en la família (v. figura 1). Per això, el gen és de naturalesa autosòmica i dominant, la qual cosa s’adiu amb la seua incidència en el 50% de la població familiar i amb la seua distribució equitativa entre homes i dones. |
Però partint de premisses menys ambicioses, hi ha alguna prova que mostre clarament una base genètica per al llenguatge? Donades les limitacions en l’estudi cerebral, la investigació d’aquesta qüestió, com succeeix amb molts altres estudis per a entendre la localització i les funcions dels gens, se centra a localitzar deficiències hereditàries que impliquen fallades en el mecanisme de transmissió del llenguatge. La lògica és clara: “Si la nostra facultat lingüística té un component innat, llavors aquest tret ha d’estar sotmès a la variació genètica” (Maynard Smith, Szathmáry, 1999: 246). I per a apreciar-ne els efectes, és necessari que l’esmentada deficiència siga específica del llenguatge i independent d’altres aspectes de la cognició.
Aquesta evidència existeix i es relaciona amb el fenomen conegut com a trastorn específic del llenguatge. Gopnik (1990: 715) va estudiar tres generacions d’una família anglesa, denominada KE, en què alguns dels seus membres (16 de 31) mostraven problemes amb el llenguatge de caràcter específicament gramatical, és a dir, que no disposen de regles generals sobre trets gramaticals (plural, temps passat, gènere, etc.). Per exemple, produeixen plurals correctes per a paraules conegudes, però manquen d’una regla general de formació de plural capaç d’operar amb paraules inventades o no conegudes: no poden decidir si el plural d’una paraula inventada, com ara wug, és wugs, i han d’aprendre la forma plural com una peça lèxica diferent en cada cas.
L’estudi posterior de famílies d’altres països va revelar que, encara que el trastorn és en part heterogeni, hi ha un nucli comú, conformat per 1) dificultats per a adquirir el llenguatge en absència de factors susceptibles de ser-ne els responsables (sordesa, retard cognitiu, dany neural aparent o autisme); 2) problemes motors (moviments orofacials) i específicament gramaticals; mentre que els primers són reversibles, i es poden superar amb el temps, els segons persisteixen. I, finalment 3), incapacitat per a usar les regles productives de formació de paraules i per a satisfer tot tipus de requisits de concordança. La singularitat del trastorn, doncs, consisteix en el caràcter específicament gramatical dels seus símptomes, així com en la seua distribució entre els membres de les famílies afectades (v. figura 1).
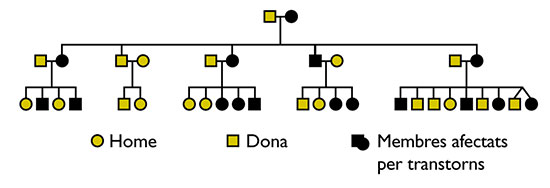
Figura 1. Arbre familiar de tres generacions de la família KE. Font: Gopnik et al., 1997: 126
Gopnik va atribuir els trastorns lingüístics de la família KE a l’herència; en concret, a un al·lel autosòmic dominant. D’altra banda, la suposició que el defecte es relaciona amb un sol gen, no amb diversos, deriva del fet que la síndrome no es produeix en graus.
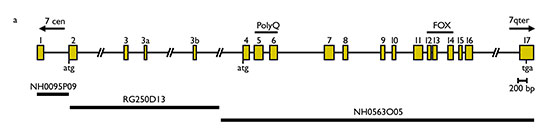
Figura 2. Representació de l’estructura del gen FOXP2. La substitució de guanina per adenina produïda en els individus afectats de la família KE es localitza en l’exó 14. Font: Lai et al., 2001: 520
La vinculació del cas descrit amb un gen es va demostrar posteriorment, en dos passos successius. En 1998, un article d’un equip de genetistes (Fisher et al., 1998: 168-170) exposa els resultats d’una recerca efectuada en el material genètic d’aquesta família. Hi va detectar una regió en el cromosoma 7, SPCH1, que en els membres afectats presenta una composició diferent de la que regeix tant en els no afectats com en la resta de la població. Aquest resultat es va concretar encara més en Lai et al. (2001: 519-523), que localitza el defecte de la família KE en un gen de la regió SPCH1, batejat com FOXP2, el primer específicament vinculat amb el llenguatge. Aquesta localització va ser corroborada mitjançant l’examen d’un altre individu (CS) sense relació amb la família; d’altra banda, anàlisis genètiques van descartar la coincidència casual o la possibilitat d’un polimorfisme natural: en tots els membres afectats de la família (a més de CS) –però en cap dels no afectats i en cap dels 364 cromosomes analitzats de persones no afectades sense relació amb la família–, un nucleòtid de guanina apareix reemplaçat per un d’adenina, la qual cosa causa un canvi d’aminoàcids (l’arginina és substituïda per l’histidina) que altera la funció de la proteïna (v. figures 2 i 3). Els autors defensen que l’alteració en aquest gen, de tipus regulador, produeix un desenvolupament anormal en l’embriogènesi de les estructures neurals rellevants per al llenguatge.

Figura 3. Representació de les tres proteïnes conegudes FOXP (1, 2 i 3), al costat de les quals es reflecteixen proteïnes d’altres branques de la família FOX. La fletxa vertical petita (damunt d’Hèlix 3) indica el lloc exacte de la substitució en FOXP2 de l’arginina per l‘histidina en els membres afectats de la família KE. Font: Lai et al. 2001: 521
El significat del descobriment
Siga la que siga la seua funció exacta, la troballa d’un gen relacionat amb el llenguatge té gran transcendència: d’una banda, mostra la factibilitat d’una genètica del comportament, que no implica determinisme, sinó aprenentatge “preparat”. D’una altra, confirma el que dicta el sentit comú: tant el nostre sistema comunicatiu, com, des de bases més àmplies, la nostra espècie, no són pas una ‘illa cultural’ situada al mig d’un mar d’instints, sinó que participem de la resta d’espècies en tant que formem part de la mateixa naturalesa. Per això, el camí obert mitjançant el descobriment és enormement prometedor, si bé ha de valorar-se en els seus justos termes. En primer lloc, de segur que hi ha altres segments genètics relacionats amb el llenguatge però no coneguts i potser no tan fàcils de localitzar, tenint en compte fenòmens com la pleitropia o efectes derivats de poligens. En tot cas, no cal suposar una gran càrrega genètica per al llenguatge, donada la rellevància dels processos d’epigènesi. D’altra banda, ha de recordar-se que les instruccions contingudes en els gens són, estrictament parlant, instruccions per a l’especialització cel·lular en tipus particulars de teixits i no poden plasmar directament principis funcionals. Per això, estudis com l’esmentat s’ha d’interpretar que confirmen l’existència de gens que codifiquen per a elaborar teixit neural específicament dedicat al llenguatge. Els anys venidors seran, de segur, apassionants en aquest sentit.
BIBLIOGRAFIA
Baker, M., 1996. The polysynthesis parameter. Oxford University Press. Nova York.
Fisher, S., Vargha-Khadem, F.,Watkins, K., Monaco, A. i M. Pembrey, 1998. “Localisation of a gene implicated in a severe speech and language disorder”, Nature Genetics, 18.
Gopnik, M., 1990. “Feature-blind grammar and dysphasia”, Nature, 344.
Gopnik, M., Dalalakis, J. i S. Fukuda, 1997. “The biological basis of language: Familial language impairment”. In Gopnik, M. [ed.]. The inheritance and innateness of grammars. Oxford University Press. Nova York.
Jackendoff, R., 2002. Foundations of language. Brain, meaning, grammar, evolution. Oxford University Press. Nova York.
Lai, C.S.L., Fisher, S. E., Hurst, J. A., Vargha-Khadem, F. i A. P. Monaco, 2001. “A forkhead domain gene is mutated in a severe speech and language disorder”, Nature, 413.
López García, A., 2002. Fundamentos genéticos del lenguaje. Cátedra. Madrid.
Lorenzo, G. i V. M. Longa, 2003. Homo Loquens. Biología y evolución del lenguaje. TrisTram. Lugo.
Maynard Smith, J. i E. Szathmáry, 1999. The origins of life. From the birth of life to the origin of language. Oxford University Press. Oxford. [Trad. de J. ROS, 2001. Ocho hitos de la evolución. Del origen de la vida a la aparición del lenguaje. Tusquets. Barcelona].
Monod, J., 1970. Le hazard et la nécessité. Du Seuil. París. [1993. El azar y la necesidad. Tusquets, 5a ed. Barcelona].
Ruse, M., 1986. Taking Darwin seriously. Blackwell. Oxford. [1994. Tomándose a Darwin en serio. Implicaciones filosóficas del darwinismo. Salvat. Barcelona].
Wilson, E. O., 1978. On human nature. Harvard University Press. Cambridge, MA. [1980. Sobre la naturaleza humana. FCE. Madrid].