|
Figura 1. Sexe i mort són dues cares d’una mateixa moneda. La teoria considera l’envelliment com una característica que ha evolucionat amb el soma. A la fotografia, hemípters de la subfamília Lygaeinae copulant. “No vull passar a la immortalitat per les meues obres, sinó per no morir-me.” La biologia evolutiva té una gran afinitat amb el pensament i les obres de Woody Allen. No en va els temes habituals d’evolucionistes i de les pel·lícules del famós director són comuns: el sexe, la mort i la religió. I és la mort o, més precisament, les seues etapes prèvies, l’envelliment, l’objecte d’aquest monogràfic, i l’aportació de la teoria evolutiva a la comprensió d’aquest el tema d’aquesta contribució. Podríem començar preguntant-nos quin sentit pot tenir analitzar l’envelliment des d’una perspectiva evolutiva si, al capdavall, l’envelliment no és sinó una etapa més de la història de la vida de tots els organismes. No obstant això, la realitat dista de ser tan simple: no tots els organismes envelleixen, ni tan sols tots moren per causes naturals, intrínseques, sense que hi intervinga l’acció d’agents externs, siguen físics, químics o biològics. Per exemple, ni els bacteris, ni les meduses i corals presenten cap dels signes que ens permeten definir l’envelliment de forma comuna per a (quasi) tots els éssers vius: una deterioració de les capacitats fisiològiques internes que es tradueix en una disminució de la probabilitat de supervivència i de la capacitat de deixar descendència i que culmina amb l’extinció dels processos bioquímics i metabòlics que caracteritzen la matèria viva. Entre els components de deterioració considerats per la teoria evolutiva queden excloses causes externes que no tinguen origen en un declinar orgànic intern. Per què uns organismes manifesten aquests senyals d’envelliment i altres no? Al cap i a la fi, el mandat bíblic “creixeu i multipliqueu-vos” no fa menció ni a envellir ni a la mort i aquests no estan necessàriament lligats a aquells que, d’altra banda i sense necessitat d’invocar creences religioses, sí que són comuns a tots els éssers vius. Història natural de l’envelliment La història natural de l’envelliment s’inicia en un període no ben definit fa entre 600 i 1.000 milions d’anys. Fins aquell moment, durant més de 2.000 milions d’anys, la Terra va estar habitada per organismes unicel·lulars, primer bacteris i arquees (caracteritzades per no disposar el seu material hereditari, el seu DNA, en un nucli separat de la resta del material cel·lular), posteriorment, també eucariotes unicel·lulars (amb nucli diferenciat), semblants a les amebes i paramecis actuals. L’èxit d’aquests organismes és innegable si es mesura per paràmetres com la persistència evolutiva de centenars i fins i tot milers de milions d’anys, la capacitat de colonització de nínxols (des de les fumaroles de les profunditats oceàniques fins a les neus perpètues), la seua capacitat de generació de noves espècies, o per la fracció de la biomassa total que representen, pròxima al 50%. Tots ells comparteixen un mateix sistema de reproducció, la reproducció asexual per fissió cel·lular, en què una cèl·lula mare es divideix i dóna lloc a dues cèl·lules filles amb què comença una nova generació. El per què –i especialment el com– aquest sistema de reproducció va ser substituït pel sexe, per la unió de dues cèl·lules procedents habitualment de distints progenitors per a donar lloc a una nova generació, representa un altre dels grans misteris de la biologia. El cas és que en el precambrià, una sèrie de protistes, que fins llavors havien avançat fins a constituir-se en organismes multicel·lulars per agregació i sense diferenciació de les cèl·lules components, comencen a posar en marxa un procés d’especialització i diferenciació cel·lular que va conduir amb el temps als fenòmens de la sexualitat, d’una banda, i la senescència o envelliment, de l’altra. Sexe i mort són dues cares d’una mateixa moneda (figura 1). El procés que porta a l’aparició de la sexualitat i la senescència és la separació entre línies cel·lulars germinals, encarregades de la formació dels gàmetes i, per tant, de la reproducció, i els llinatges cel·lulars somàtics encarregats de facilitar totes les tasques i funcions necessàries per a assegurar aquesta funció. L’especialització és escassa i incipient en les primeres etapes, però poc temps després permet la gran explosió de diversitat animal que marca l’arribada del cambrià. Aquestes línies somàtiques donen lloc a teixits i òrgans, a l’aparició de funcions especialitzades i a la possibilitat d’adaptació a condicions ambientals canviants, cosa que facilita la colonització de nous hàbitats. Però el preu de l’especialització és la pèrdua de la capacitat infinita de divisió, implícita en la reproducció asexual abans mencionada, i restringida a algunes cèl·lules de la línia germinal en les espècies amb reproducció sexual. El límit en el nombre de divisions de què és capaç una cèl·lula somàtica marca una nova propietat dels éssers vius: l’envelliment. Teoria evolutiva de l’envelliment La teoria evolutiva de l’envelliment va ser plantejada en el seu format modern per P. Medawar i G. C. Williams cap a mitjan segle XX i formalitzada en el context teòric de la genètica de poblacions per W. D. Hamilton i B. Charlesworth, entre 1966 i 1980, si bé se’n troben precedents més o menys directes en treballs d’A. Weissman i A. R. Wallace, en el segle XIX i de R. A. Fisher i J. B. S. Haldane, en la primera meitat del XX. Per exemple, Weismann, gran teòric alemany de la biologia del segle XIX, va ser el primer a separar els enfocaments fisiològic i evolutiu del problema de l’envelliment: Les consideracions fisiològiques no poden, per si soles, determinar la durada de la vida… [al contrari] la durada de la vida depèn realment de l’adaptació a condicions externes, és a dir, la seua longitud, llarga o curta, és governada per les necessitats de l’espècie. En la seua formulació més senzilla la teoria evolutiva de l’envelliment explica per què augmenta la mortalitat amb l’edat a mesura que salut i funcionalitat entren en declivi. Segons aquesta teoria, a mesura que els individus avancen en edat, la seua prolongada supervivència contribueix cada vegada menys a l’eficàcia reproductiva, perquè queda menys de la seua fertilitat total (figura 2). Com a conseqüència, la selecció natural actua menys intensament per a reduir la mortalitat en les edats avançades. La demostració formal, d’aquest postulat, present ja en els primers plantejaments evolucionistes cap a la dècada de 1880, va haver d’esperar al tractament matemàtic de Charlesworth, qui, el 1980, va demostrar que, donada l’existència de poblacions en què s’estableix una estructura d’edats i en què es presenta variació genètica per als paràmetres que les caracteritzen, l’envelliment és un corol·lari directe de la teoria de la genètica de poblacions.
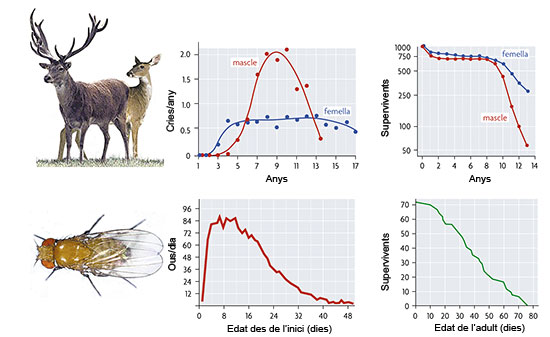
Figura 2. El procés d’envelliment es mostra com una reducció en diversos components de l’eficàcia biològica dels individus al llarg de la vida. En les figures es mostra la variació en dos d’aquests components, fecunditat i probabilitat de supervivència, en funció de l’edat en cèrvids (a dalt) i en la mosca del vinagre (a sota). Però una cosa és demostrar la correcció formal d’una teoria i una altra demostrar que siga correcta. Això ha de comprovar-se empíricament, i des de 1980 són nombrosos els estudis que han permès corroborar-la. Molts d’aquests estudis han demostrat tant l’existència de variació genètica per als diferents paràmetres descrits en la teoria plantejada per Charlesworth com la correspondència entre les seues prediccions i els resultats de diferents manipulacions experimentals. L’organisme favorit dels biòlegs que treballen en aquesta àrea és Drosophila melanogaster, la mosca del vinagre, acompanyada d’altres organismes com el escarabat de la farina, Tribolium castaneum i alguns rotífers. El plantejament general de la teoria evolutiva de l’envelliment deixa obertes nombroses qüestions, moltes de les quals amb gran interès pràctic. Entre aquestes destaca l’estudi dels mecanismes genètics que porten a l’envelliment. Aquests mecanismes solen agrupar-se en dues categories que, sense ser incompatibles, ofereixen vies alternatives per a explicar un mateix fenomen. Com que la teoria està formulada en termes genètico-poblacionals, els dos mecanismes en qüestió també segueixen aquest plantejament. Així, parlem de la pleiotropia antagonista i de l’especificitat de l’acció gènica, com els mecanismes bàsics invocats per la teoria evolutiva i que passem a explicar amb més detall a continuació. Per pleiotropia antagonista s’entén l’expressió de variants genètiques (al·lels d’un gen) que afecten diversos caràcters, de manera positiva en les primeres etapes de la vida i de manera negativa en les posteriors. No és més que la formulació genètico-poblacional d’una idea molt difosa en el pensament evolutiu, la de les restriccions o compensacions, present ja en els escrits de Darwin. G. C. Williams va ser un dels més fervents defensors d’aquest mecanisme, com veiem en l’argument següent: La selecció d’un gen que confereix un avantatge a certa edat i un desavantatge en una altra dependrà no sols de la magnitud dels seus efectes, sinó del temps en què aquests es produesquen. Un avantatge durant el període de màxima probabilitat reproductiva augmentarà la probabilitat de reproducció total més que el que la disminuirà un desavantatge proporcionalment semblant que actue posteriorment. D’aquesta manera la selecció natural freqüentment maximitzarà el vigor durant la joventut a costa del vigor en etapes posteriors, i això produirà un declivi [en l’envelliment] durant la vida adulta. [Williams, 1957] Distints estudis teòrics demostren la validesa d’aquest mecanisme per a mantenir variabilitat genètica abundant en els caràcters rellevants en condicions naturals, el que facilita, en principi, la seua validació per mitjà de l’estudi de poblacions silvestres. Hi ha dos tipus d’evidència experimental que poden indicar l’acció de mecanismes de pleiotropia antagonista: l’existència de correlacions negatives entre caràcters primerencs i tardans en la història vital i respostes contraposades a la selecció sobre els uns i els altres tipus de caràcters. El primer d’aquests ha estat demostrat en diverses espècies de Drosophila i en Caenorhabditis, on un mutant del gen age-1 confereix un augment apreciable en la longevitat a costa d’una important reducció en la fecunditat. Aquest patró es repeteix en experiments semblants amb altres espècies, no sols amb variació genètica, sinó, per exemple, en experiments de prolongació de la durada de la vida per mitjà d’alteracions de la dieta que van acompanyades d’una disminució de la fecunditat que fins i tot pot ser total. També es disposa d’exemples amb Drosophila que verifiquen que la selecció sobre la major longevitat, per exemple, comporta una disminució de la fecunditat en les edats primerenques de les mosques (figura 3). En conseqüència, la pleiotropia antagonista és un mecanisme genètic plausible per a explicar l’envelliment segons la teoria evolutiva i que té certa validació experimental, sense que açò ens permeta excloure’n l’acció d’altres mecanismes alternatius o complementaris.
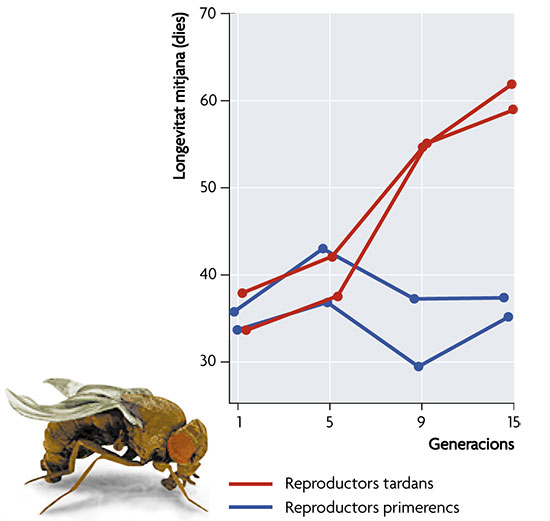
Figura 3. La gràfica mostra com la selecció artificial sobre l’etapa de la vida en què té lloc la reproducció en 4 poblacions de la mosca de la fruita, Drosophila melanogaster, dóna com a resultat un canvi en la longevitat mitjana de la població. El retard en l’acció de la selecció ve acompanyat d’una major durada de la vida. La situació respecte al segon mecanisme genètic proposat és molt semblant. Se solen reconèixer tres mètodes diferents d’acció gènica específica de l’edat que afecten l’evolució de l’envelliment: 1) la constància (estasi) de l’adaptació tardana; 2) el retard de les malalties genètiques; i 3) l’acumulació de mutacions. Considerem cada una per separat. W. D. Hamilton va proposar ja el 1996 que, atès que la intensitat de la selecció és major com menor és l’edat dels organismes, els caràcters que es manifesten a aquestes edats estarien més millorats, en una escala temporal evolutiva, que els que es manifesten a edats tardanes. Com que aquests no podrien millorar al mateix ritme, els individus més vells no podrien “mantenir el ritme” en la seua lluita evolutiva amb altres individus. Aquest mecanisme generaria un envelliment de tipus “relatiu”, en què els caràcters d’expressió tardana representen formes ancestrals no millorades per l’acció de la selecció natural. Aquesta alternativa planteja un greu problema teòric: ignora la pressió de mutació, que portaria a una degeneració encara major de les variants genètiques expressades en les etapes tardanes de la vida. Per això no pot explicar adequadament l’extrema debilitat d’individus longeus que no sofreixen els efectes de la malaltia, la prelació o la competència en el laboratori. Tant Haldane com Medawar van proposar en la dècada dels quaranta del segle passat que l’envelliment sorgeix com a conseqüència de l’extensió d’al·lels en loci modificadors que posposen l’edat de començament de greus malalties generades per altres gens. La teoria sosté que, pel fet que els efectes que s’expressen tard en la vida també tenen un menor impacte en l’eficàcia biològica, els loci modificadors que posposen els efectes deleteris actuen com un escut protector dels efectes mutacionals. Aquest mecanisme presenta alguns problemes teòrics, com ara que la selecció sobre aquests loci modificadors ha de ser molt dèbil perquè puga retardar l’edat d’inici de les malalties genètiques, però és possible comprovar-la experimentalment en els dos organismes més adequats per al seu contrast, nosaltres mateixos i les mosques Drosophila. Si la teoria és correcta, esperaríem trobar un gran nombre de trastorns genètics associats amb l’envelliment i manca d’aquests trastorns expressats en edats primerenques. Cal assenyalar que les malalties genètiques en qüestió han de ser d’etiologia genètica i específiques de l’edat, no simplement malalties associades amb l’envelliment. L’evidència de què disposem no sembla abonar aquesta teoria, perquè són abundants les malalties genètiques que s’inicien de manera congènita o en edats primerenques, com la fenilcetonúria, l’hemofília, la síndrome de Tay-Sachs, i tantes altres, mentre que escassegen les que s’inicien tard en la vida, com la malaltia de Huntington o l’hemocromatosi idiopàtica. En resum, el retard en les malalties genètiques no sembla haver tingut importància en l’evolució de la senescència. El tercer dels mecanismes d’aquest grup, l’acumulació de mutacions, també va ser suggerit per Medawar ja el 1952. Com que la intensitat amb què actua la selecció disminueix amb l’edat, la seua capacitat d’eliminar mutacions amb efectes negatius es veu cada vegada més limitada. D’aquesta forma, la senescència sorgiria com a resultat de l’acumulació de mutacions deletèries amb efectes exclusivament tardans, sempre que es mantinguen les característiques normals en les etapes primerenques en què la selecció natural és més efectiva. Les etapes més tardanes de la vida dels organismes actuarien com autèntics “poals de fem”, en què s’acumulen els al·lels amb efecte deleteri. És important destacar que aquest mecanisme ha de distingir-se de la deterioració mutacional progressiva que es produeix en la línia somàtica, com a conseqüència de l’acumulació de mutacions no reparades al llarg de la vida d’un organisme. En aquest cas, el mecanisme es basa en les mutacions que són transmissibles d’una generació a la següent, i que per tant apareixen en la línia germinal, i el principal efecte de la qual es produeix en les edats més avançades, per la qual cosa la selecció natural no és efectiva en la seua eliminació. De nou tenim una situació perfectament plausible en teoria que es pot sotmetre a l’escrutini experimental, si bé els dissenys i les interpretacions dels resultats han de ser extremadament acurats per a eludir les dificultats que planteja l’estudi en el laboratori d’organismes transferits des de la naturalesa. No obstant això, hi ha diversos experiments, novament amb Drosophila, que abonen aquesta teoria, en especial aquells que es basen en la selecció de caràcters relacionats amb un retard de la senescència i que analitzen el comportament de les poblacions en caràcters d’acció primerenca una vegada cessa la pressió de la selecció. Altres dissenys alternatius ofereixen resultats contradictoris, però, en general, hi ha un relatiu consens a considerar que l’acumulació de mutacions ofereix un mecanisme vàlid per a la teoria evolutiva de la senescència. En conjunt, la millor evidència sobre els mecanismes genètics implicats per la teoria evolutiva de la senescència prové de distints experiments amb Drosophila, encara que hi ha alguns indicadors positius addicionals en altres espècies. Tant la pleiotropia antagonista com l’acció gènica específica de l’edat, en concret l’acumulació de mutacions, poden explicar l’evolució de la senescència, però no disposem encara d’una avaluació sobre la seua importància relativa ni en el conjunt de l’evolució ni sobre espècies concretes. A falta de validació experimental directa, excepte en els casos adés mencionats, la teoria evolutiva de la senescència pot rebre (o restar) suports per mitjà de l’aplicació del mètode comparatiu. En concret, és possible aplicar aquesta metodologia tant per explicar els patrons de senescència comparant unes espècies amb altres com per contrastar aquests patrons. Aquests estudis permeten, per exemple, verificar una predicció crítica de la teoria evolutiva de la senescència, derivada de la diferenciació entre soma i germoplasma: aquelles espècies en què la línia somàtica i la línia germinal no estan clarament diferenciades no presentaran els patrons característics de l’envelliment. La teoria considera l’envelliment com una característica que ha evolucionat amb el soma i que no es presenta en la seua absència. Aquesta és una predicció fàcilment comprovable i, de fet, és una de les prediccions més fortes en biologia evolutiva. No obstant això, hi ha alguns organismes en què la separació entre soma i germoplasma no està ben definida i aquesta ambigüitat comporta dificultats afegides. Una altra font de problemes per a contrastar aquesta predicció consisteix en la diferenciació entre senescència i declivi de l’eficàcia per causes extrínseques, o intrínseques però diferents a les pròpies d’aquella (per exemple, l’acumulació de mutacions somàtiques) quan s’estudien poblacions naturals. Per a evitar-los, és necessari detectar l’envelliment com una deterioració progressiva en els components de l’eficàcia biològica específics de l’edat. Finalment, és necessari considerar la complicació dels cicles de vida que impliquen alternança entre reproducció sexual i asexual, especialment en aquells casos en què hi ha certa flexibilitat en el nombre de generacions que transcorren amb l’una i l’altra. Si no s’arriben a donar les condicions perquè un organisme entre en la fase sexual del seu cicle reproductiu, pot observar-se una disminució progressiva del seu vigor, sense que açò puga interpretar-se com un cas de senescència, sinó de falta de reproducció. Amb totes aquestes consideracions en ment, l’evidència disponible, des de vertebrats a protozous, passant per plantes i invertebrats, és prou clara: no hi ha casos en la literatura comparativa que refuten amb claredat la teoria evolutiva de l’envelliment. A banda de la predicció forta mencionada adés, la teoria evolutiva proporciona altres prediccions de menor rang susceptibles de comprovació per mètodes comparatius. Entre les més rellevants es troben els següents corol·laris de la teoria plantejats per Williams (1957): 1. Les baixes (altes) taxes de mortalitat adulta haurien d’estar associades amb baixes (altes) taxes d’envelliment. 2. L’envelliment ha de ser més ràpid en aquells organismes que no augmenten substancialment la fecunditat després de la maduresa que en aquells que sí que presenten aquest increment. 3. Sempre que hi haja diferències entre els sexes, el sexe amb major taxa de mortalitat i menor taxa d’augment de la fecunditat hauria d’experimentar un envelliment més ràpid. 4. La reproducció primerenca s’ha de correlacionar amb un envelliment primerenc i, recíprocament, els organismes més longeus han de presentar taxes de reproducció primerenca més baixes. Aquest últim corol·lari es deriva del mecanisme de pleiotropia antagonista exposat adés i no de la teoria general evolutiva de l’envelliment. Què ens diu l’evidència experimental disponible? En general, les dades abonen les quatre prediccions anteriors, però es presenten excepcions que encara no han estat acomodades en el marc teòric. La teoria evolutiva ofereix per a l’envelliment una explicació raonable, ben integrada en el seu marc teòric i amb suficients evidències empíriques. S’hi estableix que l’evolució de l’envelliment cal trobar-la en la disminució progressiva amb l’edat de l’eficàcia de la selecció natural en aquells organismes que tenen reproducció sexual i en què hi ha una clara diferenciació entre soma i germoplasma. En aquest marc general, la teoria exposada no té rival en l’actualitat, però, a mesura que descendim en els nivells explicatius i busquem mecanismes que li permeten actuar efectivament, el que trobem és una pluralitat d’alternatives, amb nivells variables de plausibilitat i validació experimental, cap de les quals aconsegueix, en el seu nivell, explicar de forma necessària i suficient per si sola la varietat de processos fisiològics i evolutius implicats en l’envelliment. Per tant, hem d’acceptar que la pluralitat de mecanismes proposats també ha de correspondre, en una mesura encara per determinar, amb una varietat de causes operatives, sempre que no entren en col·lisió les unes amb les altres ni, per descomptat, amb la teoria general plantejada. Aquesta pluralitat de mecanismes és una font constant de controvèrsia, però també d’estímul per a la investigació. Bibliografia Fernando González Candelas. Institut Cavanilles de Biodiversitat i Biologia Evolutiva i Departament de Genètica, Universitat de València. |
«No tots els organismes envelleixen, ni tan sols tots moren per causes naturals, intrínseques, sense que hi intervinga l’acció d’agents externs»
«L’evolució de l’envelliment cal trobar-la en la disminució progressiva amb l’edat de l’eficàcia de la selecció natural en aquells organismes que tenen reproducció sexual i en què hi ha una clara diferenciació entre soma i germoplasma»
|
Per què ens fem vells? Una visió evolutiva