Moviments de dispersió en els primats
Variabilitat en els seus patrons i causes

Els moviments de dispersió en els animals representen decisions crucials per als individus, ja que afecten la seua supervivència i èxit reproductiu, a més de ser un component important de la dinàmica poblacional. En aquest article es descriu la variabilitat en els patrons de dispersió en els primats i algunes de les causes, tant últimes com proximals, a les quals respon.
Paraules clau: competència intragrupal, elecció de parella, infanticidi, depressió consanguínia.
«La dispersió proporciona una connexió genètica entre les poblacions i influeix de manera important en els processos demogràfics i en la dinàmica poblacional»
La distribució espacial dels organismes no sol ser aleatòria, sinó que respon als factors biòtics i abiòtics presents en l’ambient. Aquesta distribució, ja siga agregada o dispersa, és el resultat del moviment dels individus cap a la localització que ocupen en cada moment. En aquest sentit, els conceptes de migració i dispersió s’utilitzen per a descriure, de manera general, les característiques de certs moviments que realitzen els organismes. La migració fa referència al moviment conjunt d’un nombre elevat d’individus de la mateixa espècie entre dos hàbitats distints. Els moviments migratoris poden ser cíclics (amb distinta periodicitat, per exemple diària o anual) o d’anada únicament, la qual cosa dóna lloc a diversos patrons migratoris. La dispersió fa referència al moviment que provoca l’allunyament dels individus de les seues àrees natals i/o dels seus progenitors i individus pròxims. Aquests moviments es poden produir en solitari o en petits grups i poden ser actius o passius. La dispersió és una de les característiques clau de les poblacions pel fet que proporciona una connexió genètica entre aquestes i influeix de manera important en els processos demogràfics i en la dinàmica poblacional. En aquest article ens centrarem en els moviments de dispersió en els primats.
Dispersió en els primats
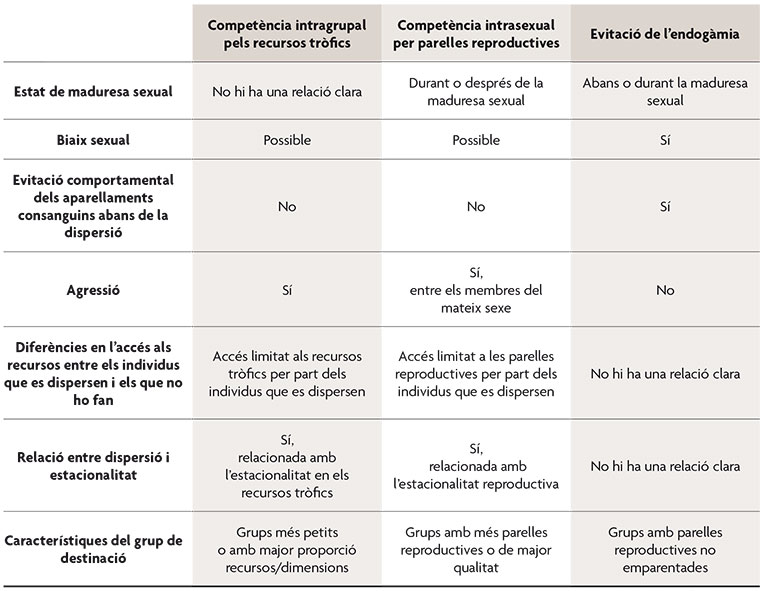
Taula 1. Característiques dels patrons de dispersió i la possible relació que mantenen amb les tres principals causes últimes de la dispersió natal. / Adaptat de la taula 2 de Field i Guatelli-Steinberg, 2003.
Dispersió en els primats
L’ordre dels primats, al qual pertany l’espècie humana, és constituït per espècies altament socials els individus de les quals viuen en grups, en molts casos durant tota la vida. En aquest sentit la dispersió dels primats, i en general de les espècies socials, té dues facetes: l’ecològica (dispersió espacial) i la social (dispersió social) (Isbell i Van Vuren, 1996). En la dispersió espacial els individus es traslladen a àrees domèstiques (home range) desconegudes, mentre que en la dispersió social els individus es traslladen a nous grups socials. Aquests dos aspectes de la dispersió es poden produir de manera conjunta o no, depenent, principalment, del tipus d’organització social dels grups. En aquelles espècies els grups de les quals defensen un territori, com en el cas dels ximpanzés, la dispersió dels individus sol combinar ambdós aspectes. En altres espècies, com els goril·les o els papions sagrats, les àrees domèstiques dels grups socials se solapen, per la qual cosa, generalment, la dispersió dels individus posseeix un component social però no ecològic. Finalment, la dispersió espacial en absència de dispersió social implicaria el desplaçament de tot el grup social a una àrea domèstica diferent. Aquests desplaçaments solen ser el resultat d’alteracions en l’hàbitat que afecten recursos o factors vitals per a l’espècie, la qual cosa impulsa el grup a cercar nous llocs amb condicions més adients. Encara que hem considerat aquest tipus de moviment com un cas de dispersió espacial, també podria ser valorat com un procés migratori no estacional.
Per què abandonar l’entorn natal o conegut?
La incorporació a un nou grup social representa en molts casos la pèrdua de les relacions socials cooperatives establides en l’anterior grup i la necessitat d’establir noves relacions, amb la consegüent despesa en temps i en energia. Tot això té lloc en un entorn social generalment hostil en el qual els membres del grup (o un subgrup d’aquest) agredeixen l’individu immigrant per desplaçar-lo dels llocs d’alimentació i evitar que s’incorpore al grup. A més, si la dispersió té lloc cap a una àrea desconeguda, l’individu pot esdevenir més vulnerable a l’atac dels depredadors i patir certs costos com els derivats del desconeixement de la localització dels recursos alimentaris. Encara que hi ha estratègies per a reduir aquests costos, com la dispersió en companyia de familiars o l’establiment d’algunes relacions amb el grup abans d’incorporar-s’hi, la dispersió és clarament un procés costós per als individus implicats, la qual cosa ens porta a plantejar-nos per què els individus exhibeixen aquest comportament.
«La incorporació a un nou grup social representa en molts casos la pèrdua de les relacions socials cooperatives establides en l’anterior grup»
A l’hora d’intentar respondre aquesta qüestió cal distingir entre les causes proximals, que fan referència als mecanismes implicats en l’expressió del comportament, i les causes últimes, que estan relacionades amb les conseqüències adaptatives del comportament i, per tant, amb l’explicació de la selecció i evolució. Entre les possibles causes proximals per a la dispersió en primats es troben: l’atracció per individus d’altres grups, els factors fisiològics, l’estat nutricional de l’individu, les taxes d’interacció amb els membres del grup (entre les quals s’inclouen les agressions o les interaccions sexuals), la coerció o la resposta a pertorbacions ambientals. En relació a les causes últimes de la dispersió en primats, s’han suggerit quatre grans categories. En primer lloc, els individus podrien abandonar el seu grup per evitar la competència pels recursos tròfics amb els membres d’aquest, especialment amb els seus parents. En segon lloc, els individus podrien abandonar el seu grup en resposta a la competència intrasexual per les parelles reproductives, especialment la que ocorre entre parents. En tercer lloc, les femelles podrien emprar la dispersió com una estratègia contra l’infanticidi comès pels mascles. L’infanticidi representa una estratègia adaptativa per a certs mascles no emparentats amb les cries pel fet que redueix el període d’infertilitat postpart i accelera la taxa reproductiva de la femella. Finalment, els individus que abandonen el grup natal podrien obtenir un benefici si s’aparellen amb individus no emparentats i eviten els efectes negatius per a la supervivència de les cries derivats dels aparellaments endogàmics (i. e. depressió consanguínia).

En la imatge, dos langurs Hanuman (Semnopithecus entellus) a l’Índia. Aquesta espècie de cercopitècids asiàtics forma grups de 15 a 54 individus entre els quals podem trobar un sol mascle reproductor o diversos. La dispersió és masculina i està associada amb la competència per les parelles reproductives. / Daniele Colombo
Aquests quatre beneficis associats a la dispersió no són mútuament excloents, ja que, encara que la causa primària de la dispersió residira, per exemple, en l’evitació de la competència intragrupal pels recursos tròfics, en dispersar-se un dels sexes com a conseqüència d’aquesta pressió selectiva es podria, al seu torn, reduir el risc d’aparellaments endogàmics en la població o la competència per les parelles reproductives. L’estudi dels patrons de dispersió i, concretament, la determinació de quins individus i en quin moment la practiquen i del mode de dispersió ens pot ajudar a determinar les causes últimes de la dispersió en cada cas (taula 1). Al seu torn, el lloc o grup cap al qual es dispersen els individus també constitueix un factor important que es troba reflectit en alguns dels exemples que presentem en els següents apartats. D’aquesta manera, per exemple, quan la dispersió de les femelles no es troba associada amb la maduresa sexual sinó amb un increment de l’agressió intrasexual dins del grup i, al seu torn, les femelles tendeixen a desplaçar-se a grups menys densos o amb àrees domèstiques de major qualitat en termes de recursos tròfics, podem postular que la causa més probable de la dispersió és la competència pels recursos tròfics dins del grup.
Característiques dels patrons de dispersió
Qui es dispersa?
En la majoria de mamífers hi ha un caire en el patró de dispersió en relació amb el sexe: és el mascle el que generalment abandona la unitat natal. Això és degut a l’asimetria en la inversió parental que es produeix de manera general en els mamífers (les femelles hi inverteixen més que els mascles) i que afecta les taxes reproductives potencials (nombre de cries que cada sexe podria produir per unitat de temps si la seua reproducció no es trobara limitada per la disponibilitat de parelles reproductives) i els factors que limiten l’èxit reproductiu en l’un i l’altre sexe (Trivers, 1972). D’una banda, romandre en l’entorn natal permet a les femelles tenir accés a una font d’aliment coneguda, que representa el factor limitant per a la seua reproducció i, al seu torn, els facilita l’establiment d’una sèrie de relacions cooperatives entre individus emparentats (en dispersar-se els mascles, aquestes relacions s’establirien entre femelles emparentades i formarien les anomenades matrilínies) i no emparentats que té un impacte positiu sobre la seua supervivència i reproducció. D’altra banda, encara que els mascles podrien beneficiar-se igualment de romandre en el seu entorn natal, el seu èxit reproductiu es troba limitat pel nombre de parelles a què tenen accés, per la qual cosa els costos de la dispersió podrien veure’s compensats incorporant-se a unitats en què optimitzaren aquest accés.
«En la majoria de mamífers hi ha un caire en el patró de dispersió en relació amb el sexe i és el mascle el que generalment abandona la unitat natal»
En primats hi ha diverses espècies que poden incloure’s dins d’aquest patró general de dispersió (taula 2). Un exemple de dispersió masculina associada a la competència per les parelles reproductores el trobem en els langurs comuns o Hanuman (Koenig i Borries, 2001). Aquesta espècie, pertanyent a la família dels cercopitècids, pot formar grups amb només un mascle adult i diverses femelles (unimascle-multifemella) o grups multimascle-multifemella. En el primer cas, els mascles solen abandonar el grup al voltant de la maduresa sexual, encara que poden ser-ne expulsats abans si el mascle dominant (possiblement el seu pare) és reemplaçat per un altre individu aliè al grup. D’altra banda, en els grups multimascle-multifemella, els individus poden romandre durant més temps en les seues unitats natals abans de dispersar-se. En ambdós casos, la competència per les parelles reproductores es postula com la principal causa de la dispersió. En el primer cas, un mascle és capaç d’expulsar la resta de competidors de la unitat i, en l’altre, el mascle dominant és el que monopolitza la major part de la paternitat en el grup. Els mascles que es dispersen solen unir-se a grups que només contenen mascles en espera de formar o accedir a un grup reproductor.
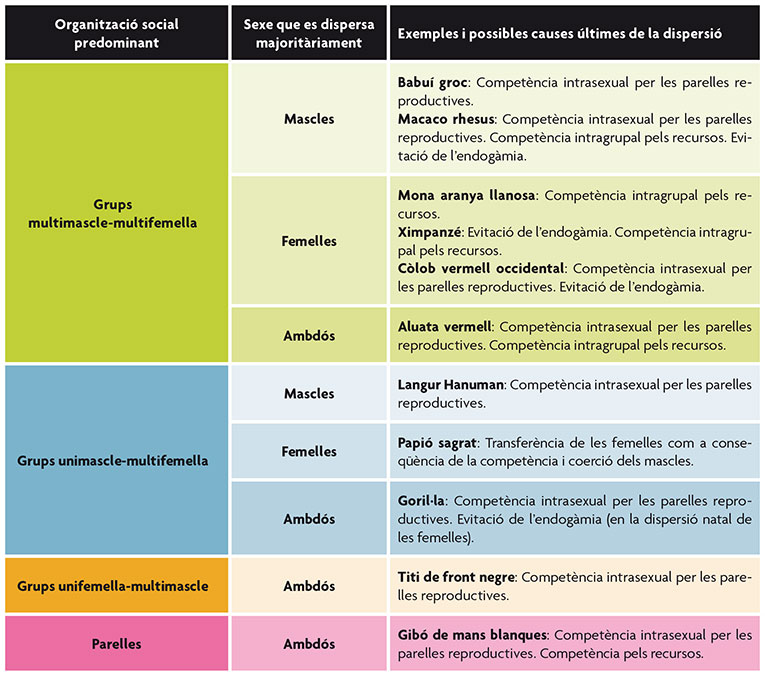
Taula 2. Principals patrons de dispersió descrits en primats en funció del tipus predominant d’organització social de l’espècie i del caire sexual en la dispersió. Al seu torn, es mostren diversos exemples i les causes últimes més probables de la dispersió en cada cas.
No obstant això, hi ha diverses espècies que no s’ajusten a aquest patró general de dispersió i en les quals les femelles habitualment abandonen les unitats natals mentre que els mascles hi romanen (taula 2). Es considera que la competència pels recursos tròfics constitueix la causa més freqüent que determina aquest patró, encara que l’evitació de l’endogàmia o la tria de parella també constitueixen causes probables en certs casos. Les mones aranya llanoses són una espècie del Nou Món, pertanyent a la família dels atèlids, que formen grups multimascle-multifemella amb un sistema de fusió-fissió en què les femelles abandonen habitualment el grup natal al voltant de la maduresa sexual. Encara que no són expulsades agressivament del grup, les femelles adultes les desplacen ben sovint dels llocs d’alimentació, per la qual cosa es pensa que aquesta dispersió és deguda en gran manera a la competència pels recursos tròfics (Printes i Strier, 1999). Els ximpanzés, pertanyents a la família dels homínids, també presenten sistemes socials de fusió-fissió formats per grups multimascle-multifemella (Stumpf, 2007). Els mascles són filopàtrics, ja que romanen en les seues comunitats natals, en les quals formen coalicions i cooperen en la defensa del territori, i són les femelles les que es dispersen a altres grups al voltant de la maduresa sexual. No obstant això, a diferència del que ocorre en el cas de les mones aranya llanoses, aquesta dispersió sembla més aviat relacionada amb l’atracció de les femelles cap als mascles de fora del grup que no amb la competició per l’aliment dins del grup. En aquest cas, les causes proximals de la dispersió semblen indicar que aquesta està majoritàriament relacionada amb l’evitació de l’endogàmia, encara que la competència pels recursos tròfics podria estar representant un cert paper en el procés. Finalment, l’espècie humana també representa una excepció al patró general de dispersió en els primats, ja que, encara que ambdós sexes poden abandonar les seues unitats natals, la dispersió es troba esbiaixada cap a les femelles en la majoria de les societats estudiades (Rodseth et al., 1991).
En altres espècies no hi ha un biaix sexual en el patró de dispersió, ja que ambdós sexes abandonen el grup natal (taula 2).
Els goril·les, pertanyents a la família dels homínids, constitueixen un exemple d’aquest patró de dispersió. Els seus grups socials són formats per un sol mascle reproductor o, de vegades, per dos, encara que el dominant acapara la majoria dels aparellaments i, per tant, de la paternitat en el grup (Harcourt i Stewart, 2007). Conseqüentment, la principal causa de dispersió en els mascles seria la competència reproductiva. En el cas de les femelles, aquesta dispersió està relacionada amb l’evitació dels aparellaments endogàmics i amb l’elecció de parella.
«L’infanticidi representa una estratègia adaptativa per a certs mascles no emparentats amb les cries perquè redueix el període d’infertilitat postpart»
Per finalitzar, cal assenyalar que el tipus de biaix sexual en la dispersió constitueix un factor important ja que determinarà l’estructura de parentiu dins dels grups i influirà sobre les característiques de les relacions socials (i. e. estructura social). En general, el sexe que es dispersa perd la major part de les relacions socials amb els seus parents. Al seu torn, els vincles que estableix en el nou grup amb els individus del mateix sexe solen ser dèbils o inexistents. En contraposició, el sexe filopàtric, el que roman en el seu entorn natal, estableix forts vincles cooperatius i aliances amb els seus parents i forma el nucli de l’estructura social del grup (matrilínies o patrilínies, depenent del sexe). D’aquesta manera, filopatria, consanguinitat i cooperació solen coincidir. En l’espècie humana, però, les femelles tendeixen a mantenir els vincles amb els seus parents, a pesar de separar-se’n, excepte en societats amb un component patriarcal extrem. Al seu torn, també poden formar vincles cooperatius en els seus nous grups amb femelles amb les quals no estan emparentades, circumstància que es dóna en menor grau en altres espècies de primats amb dispersió femenina amb excepció, potser, dels bonobos. D’altra banda, en aquelles societats humanes en què les femelles són filopàtriques, els mascles tampoc perden els vincles amb els seus parents i, de fet, aquestes societats solen estar caracteritzades per un alt grau d’endogàmia comunitària, ja que els mascles o no abandonen la comunitat natal o es desplacen a grups veïns, moltes vegades dependents de la comunitat natal (Rodseth et al., 1991).
Quan dispersar-se? El moment de la dispersió i la relació amb l’infanticidi
Podem parlar de dos tipus de dispersió social en funció del moment en què té lloc. La dispersió primària (o natal) és la que ocorre al voltant de la maduresa sexual i representa una separació de l’individu del seu grup natal, mentre que la dispersió secundària constitueix un canvi d’unitat social durant l’edat adulta.
«L’espècie humana representa una excepció al patró general de dispersió en els primats, ja que la dispersió es troba esbiaixada cap a les femelles en la majoria de les societats estudiades»
El moment en què els individus es dispersen de la seua unitat natal pot ser crític per a la seua supervivència, no obstant això, aquest és un paràmetre que varia substancialment entre espècies i per a diferents individus dins d’una mateixa espècie, com hem vist en el cas dels langurs Hanuman. Es pensa que el moment de la dispersió és determinat pel balanç entre els costos potencials de romandre en el grup (depressió consanguínia, agressió per part dels membres del grup, accés limitat als recursos tròfics i socials) i els costos potencials d’abandonar-lo, que han estat descrits prèviament.
En algunes espècies en què les femelles es dispersen secundàriament, la decisió de quan fer-ho es troba influïda pel risc d’infanticidi (Sterck i Korstjens, 2000). En primer lloc, per norma general, les femelles canvien de grup social quan no tenen cries dependents a càrrec seu per evitar agressions infanticides comeses pels mascles en els nous grups. I en segon lloc, en aquelles espècies, com els goril·les, els langurs de Thomas o els còlobs vermells, en què la paternitat es concentra en un sol mascle o hi està fortament esbiaixada, la percepció del risc que aquest mascle siga reemplaçat per un altre d’extern al grup, circumstància que ocasionaria una situació de risc d’infanticidi per a les cries, és el factor que desencadena la dispersió de les femelles no lactants, de vegades, al grup del potencial mascle infanticida. Aquesta dispersió s’interpreta des del punt de vista de l’elecció de parella, ja que les femelles sembla que canvien d’unitat social en cerca d’un mascle que els proporcione una millor protecció per a les seues cries enfront de les agressions.
El mode de dispersió. Conflicte sexual i dispersió en els papions sagrats
«La dispersió es pot produir per iniciativa pròpia o com a conseqüència del rebuig i agressió per part dels individus del grup»
Com ja hem avançat, la dispersió es pot produir per iniciativa pròpia o com a conseqüència del rebuig i agressió per part dels individus del grup. No obstant això, en els papions sagrats, pertanyents a la família dels cercopitècids, les femelles no es dispersen per cap d’aquests dos processos sinó que són «segrestades». El segrest és un tipus de coerció sexual que implica la separació forçosa d’una femella de la seua unitat social. En els papions sagrats aquest és un procés complex en què els mascles competeixen per la femella i aquesta, al seu torn, rep una agressió intensa del mascle segrestador que té com a objectiu condicionar el comportament de la femella a canviar d’unitat i té associat un cost per a l’èxit reproductiu immediat de la femella (Polo i Colmenares, 2012; Polo et al., 2014). D’aquesta manera, la dispersió de les femelles entre les unitats unimascle, que constitueix la unitat social i reproductiva bàsica en aquesta espècie, està més relacionada amb una estratègia coercitiva del mascle que no amb una estratègia reproductiva de la femella, ja que el canvi de grup és forçat pel mascle i és costós per a la femella.

En els papions sagrats (Papio hamadryas hamadryas) els mascles agredeixen amb intensitat les femelles que estan sent transferides a les seues unitats. El mos al bescoll és una de les agressions característiques que es produeix durant aquests episodis denominats pasturatges. / Pablo Polo
Consideracions finals
A mesura que els patrons de dispersió es caracteritzen millor i per a un nombre més gran d’espècies, s’observa que hi ha nombroses excepcions als patrons generals que s’han descrit. En primer lloc, encara que la causa última de la dispersió dels mascles es relaciona en moltes espècies amb la competència intragrupal per l’accés a les parelles reproductives, també hi ha casos en què aquesta causa sembla estar més relacionada amb la competència per l’aliment dins del grup o l’evitació de l’endogàmia. Al seu torn, la dispersió en les femelles sembla estar més relacionada amb la competència per l’aliment dins del grup o l’evitació de l’endogàmia, no obstant això, en certes espècies es pensa que les femelles també podrien abandonar el seu grup en cerca de millors oportunitats reproductives. De fet, segons el consens actual, es considera que la dispersió en primats apareix com a resultat de la combinació d’aquestes causes o d’algunes d’elles, tant a escala interespecífica com intraespecífica. I, en segon lloc, no tots els individus d’una població es comporten segons el patró general de dispersió d’aquesta població. Per exemple, en alguns casos, les femelles de ximpanzés no es dispersen i romanen durant tota la vida en els grups natals. Al seu torn, s’han descrit casos en espècies en què les femelles constitueixen el sexe filopàtric, en què algunes d’aquestes abandonen el seu grup natal (per exemple, en les mones caputxines). Aquests casos es poden explicar mitjançant una aproximació al fenomen de la dispersió basada en l’individu, pel fet que els costos i beneficis de la dispersió no depenen només de l’espècie i del sexe, sinó que també ho són de la situació de cada individu en el grup (Jack i Isbell, 2009). De manera que, per a certes femelles, en espècies amb filopatria femenina, el balanç entre costos i beneficis pot afavorir l’abandó del grup a causa, per exemple, d’una baixa posició en la jerarquia de dominància del grup que limitaria el seu accés als recursos. Aquestes dues característiques que hem descrit provoquen l’aparició de la gran variabilitat tant interespecífica com intraespecífica i interindividual que trobem en els patrons de dispersió en els primats.
Field, M. i D. Guatelli-Steinberg, 2003. «Dispersal and the Inbreeding Avoidance Hypothesis». Primate Report, 67: 7-60.
Harcourt, A. H. i K. J. Stewart, 2007. Gorilla Society: Compromise and Cooperation between Sexes. University of Chicago Press. Chicago.
Isbell, L. A. i D. Van Vuren, 1996. «Differential Costs of Locational and Social Dispersal and Their Consequences for Female Group-living Primates». Behaviour, 133: 1-36. DOI: <10.1163/156853996X00017>.
Jack, K. M. i L. A. Isbell, 2009. «Dispersal in Primates: Advancing An Individualized Approach». Behaviour, 146: 429-436. DOI: <10.1163/156853909X410612>.
Koenig, A. i C. Borries, 2001. «Socioecology of Hanuman Langurs: the Story of Their Success». Evolutionary Anthropology: Issues, News, and Reviews, 10: 122-137. DOI: <10.1002/evan.1026>.
Polo, P. i F. Colmenares, 2012. «Behavioural Processes in Social Context: Female Abductions, Male Herding and Female Grooming in Hamadryas Baboons». Behavioural Processes, 90: 238-245. DOI: <10.1016/j.beproc.2012.02.004>.
Polo, P.; Hernández-Lloreda, M. V. i F. Colmenares, 2014. «Male Takeovers Are Reproductively Costly to Females in Hamadryas Baboons: A Test of The Sexual Coercion Hypothesis». PLOS ONE, 9(3): e90996. DOI: <10.1371/journal.pone.0090996>.
Printes, R. C. i K. B. Strier, 1999. «Behavioral Correlates of Dispersal in Female Muriquis (Brachyteles arachnoides)». International Journal of Primatology, 20: 941-960. DOI: <10.1023/a:1020882719850>.
Rodseth, L. et al., 1991. «The Human Community as a Primate Society [and Comments]». Current Anthropology, 32: 221-254. DOI: <10.2307/2743773>.
Sterck, E. H. M. i A. H. Korstjens, 2000. «Female Dispersal and Infantice Avoidance in Primates». En Van Schaik, C. P. i C. H. Janson (eds.). Infanticide by Males and its Implications. Cambridge University Press. Cambridge.
Stumpf, R., 2007. «Chimpanzees and Bonobos: Diversity within and between Species». En Campbell, C. J.; Fuentes, A.; MacKinnon, K. C.; Panger, M. y S. K. Bearder (eds.). Primates in Perspective. Oxford University Press. Oxford.
Trivers, R., 1972. «Parental Investment and Sexual Selection». En Campbell, B. (ed.). Sexual Selection and The Descent of Man 1871-1971. Aldine-Atherton. Chicago.