

Sauròpodes virtuals
Una revolució en l’estudi dels dinosaures de coll llarg

L’estudi dels fòssils mitjançant tècniques de visualització en tres dimensions, més conegut com a paleontologia virtual, ha suposat una revolució en la investigació dels organismes del passat en els darrers vint anys. L’estudi dels populars dinosaures sauròpodes n’és un exemple: els complexos sistemes de suports o les costoses rèpliques usats antany per estudiar els seus esquelets són ara fets fàcilment gràcies als avenços en computació. Qüestions com la posició dels seus llargs colls, les seues capacitats de moviment i les seues implicacions funcionals o fins i tot la còpula són més fàcils d’abordar que mai. Així, evidències obtingudes mitjançant la metodologia tradicional juntament amb les aportacions de les noves tecnologies obren noves fronteres a l’estudi d’aquests colossos.
Paraules clau: paleontologia virtual, rang de moviment del coll, còpula, girafes, Juràssic.
Introducció: un revulsiu per a la paleontologia
Els sauròpodes eren dinosaures amb un pla corporal característic: quadrúpedes de llargues cues i colls que adquiriren enormes mides. Els més grans arribaren a assolir l’alçada d’edificis de quatre pisos i masses de fins a 70 tones (tant com 250 elefants), cosa que els converteix en els animals més grans que mai hagen caminat sobre la Terra (Hallett i Wedel, 2016; Sander et al., 2010). Com se’n sortien aquests animals tan impressionants en els seus ambients pretèrits ha estat interpretat de múltiples maneres des que els primers esquelets raonablement complets van ser descoberts a finals del segle XIX, en un debat que s’ha prolongat fins ara (Taylor, 2010). Considerats durant dècades com a animals obligatòriament aquàtics a causa de la seua gran grandària, passaren a ser interpretats a finals del segle XX com a girafes reptilianes: animals fitòfags (menjadors de plantes) amb colls imponents coronats per petits caps que s’alimentaven de les copes dels arbres (Bakker, 1971). Tot i això, l’estudi dels fòssils a partir de visualització 3D, més conegut com a paleontologia virtual, ha suposat un revulsiu no només en l’estudi dels fòssils en general (Sutton et al., 2014), sinó també en la interpretació dels dinosaures sauròpodes des de principis de la dècada dels 2000 (Stevens, 2013; Stevens i Parrish, 1999; Taylor et al., 2009; Vidal, Mocho, Aberasturi et al., 2020; Vidal, Mocho, Páramo et al., 2020).
El debat sobre la posició del coll
Els ràpids avenços en tecnologia computacional han permès obtenir informació sense precedents dels fòssils de dinosaures. En el cas dels dinosaures sauròpodes, aquesta tecnologia ens ha permès des d’estudiar l’interior de les cavitats dels seus ossos (com les del sistema nerviós central o l’oïda) –sense necessitat d’alterar-los físicament gràcies a la tomografia computeritzada– fins a una cosa tan simple però revolucionària com poder treballar en un entorn virtual simultàniament amb desenes d’ossos que en el món físic pesen centenars de quilos cadascun i resulten molt complicats de manejar.
Inicialment, aquesta tecnologia feu possible un estudi que va comparar els colls dels populars sauròpodes Apatosaurus i Diplodocus per mesurar el seu rang de moviment (Stevens i Parrish, 1999). L’estudi va digitalitzar les facetes articulars de les vèrtebres cervicals d’aquests dinosaures a partir de les il·lustracions dels ossos publicades a les descripcions monogràfiques d’aquests tàxons i va calcular quant moviment eren capaços d’exercir. Els resultats mostraren que ni Apatosaurus ni Diplodocus tenien colls tan flexibles com el d’una girafa actual, ni serien capaços d’elevar els colls molt per sobre de les espatlles (Stevens i Parrish, 1999). Estudis posteriors semblaven corroborar aquesta hipòtesi, mostrant, a més, que la columna vertebral de moltes espècies de dinosaures sauròpodes s’articulava pràcticament recta, sense una elevació del coll a la base com passa en molts dinosaures teròpodes (Stevens i Parrish, 2005).
La conclusió era clara: aquesta imatge dels dinosaures sauròpodes com a girafes reptilianes que s’alimentaven de les copes dels arbres havia de ser revisada. A més, noves troballes de fòssils com el petit Nigersaurus de fa 110 milions d’anys mostraven clares adaptacions a una alimentació de vegetació a nivell del sòl com un crani el musell del qual estava orientat cap avall, extremitats anteriors curtes o una bateria dental amb desenes de dents de reemplaçament (Sereno et al., 2007). Aquestes troballes aviat influïren en com s’interpretava la major part de sauròpodes: animals amb colls horitzontals molt poc flexibles, gairebé incapaços de mirar el que tenien a la seua esquena ni d’aixecar el coll, la qual cosa va donar com a resultat representacions com la de Diplodocus al cèlebre documental de 1999 Walking with dinosaurs de la BBC.
Tanmateix, aquesta hipòtesi tan estesa de sauròpodes de colls poc flexibles i més o menys horitzontals no fou acceptada unànimement per la comunitat científica. Es va constatar que els vertebrats terrestres actuals, des dels conills fins als camells o els cocodrils, habitualment sostenen el coll en postures més elevades del que suggerien les dades de la paleontologia virtual (Taylor et al., 2009). Vol dir això que els models virtuals no permeten inferir les capacitats dels vertebrats i que estan massa limitats?
Spinophorosaurus virtual: un nou canvi d’interpretació
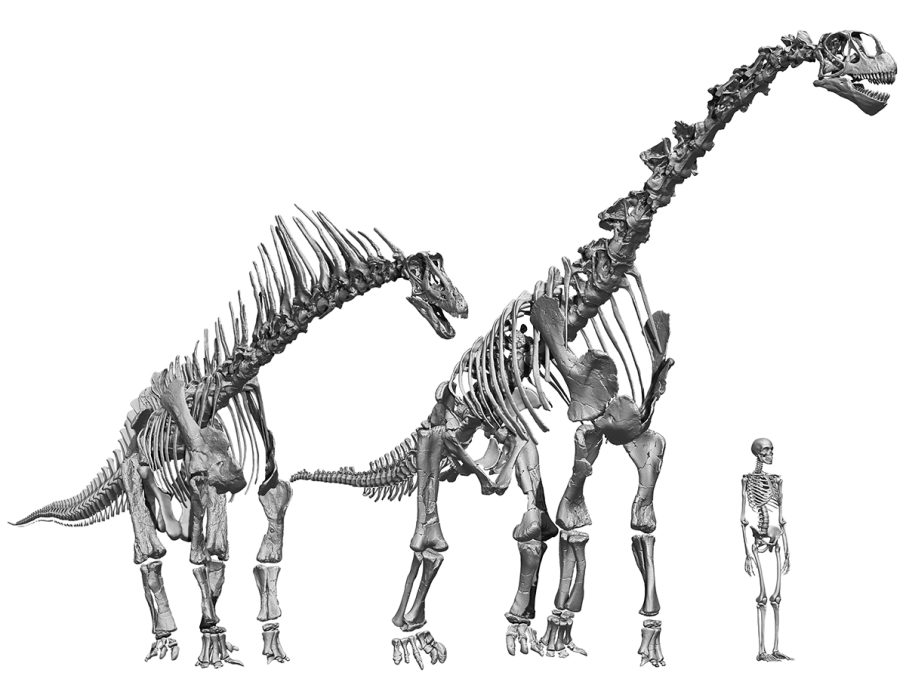
Els estudis duts a terme en Spinophorosaurus nigerensis, un sauròpode primerenc excepcionalment complet i ben preservat del període juràssic del Níger (gairebé 170 milions d’anys), van combinar ambdues línies d’evidència, les quals van mostrar que, lluny de ser incompatibles, són perfectament complementàries. Spinophorosaurus es coneix a partir de tres exemplars articulats (Remes et al., 2009): un espectacular esquelet gairebé complet i articulat d’un subadult d’11 metres de longitud, restes parcials d’un adult de 13 metres de longitud i el coll i les primeres vèrtebres de l’esquena d’un juvenil de 3 metres.
A diferència de treballs previs, on es van digitalitzar il·lustracions en dues dimensions per generar una projecció tridimensional, els ossos d’aquests tres Spinophorosaurus van ser tots digitalitzats individualment mitjançant una tècnica anomenada fotogrametria digital, per elaborar un esquelet virtual complet de Spinophorosaurus. Aquesta tècnica, basada en el mateix principi bàsic que un estereoscopi, consisteix en la presa de moltes fotografies lleugerament superposades que recullen tota la superfície de cadascun dels ossos fòssils de l’esquelet (Mallison i Wings, 2014). El seu processament en un ordinador permet obtenir moltíssims punts homòlegs entre totes les fotografies, que es fan servir per generar una superfície virtual que reprodueix el fòssil amb un marge d’error inferior al mil·límetre –si l’escaneig s’ha dut a terme correctament–. Si es repeteix aquest procés amb cada os, s’obté una col·lecció d’ossos virtuals a punt per ser articulats.
El primer pas per tenir una noció de com es disposa l’esquelet d’un vertebrat és articular els ossos en allò que s’anomena postura osteològica neutra, aquella posició que resulta d’articular els ossos en el màxim contacte que permeten les facetes d’articulació. Tot i que s’anomena neutra, això no implica necessàriament que siga la postura més habitual que adopta un animal en vida (Dzemski i Christian, 2007). Es tracta d’un punt de partida relativament objectiu que permet comparar la disposició de l’esquelet entre diferents espècies de vertebrats sota un mateix criteri: el màxim contacte articular. Intentant evitar nocions preconcebudes de quina podria ser aquesta postura osteològica neutra, la columna vertebral s’articula únicament amb una parella de vèrtebres visibles alhora. Mantenint una vèrtebra completament estàtica, se n’articula la següent amb totes les facetes d’articulació en contacte. Un cop articulades aquestes dues primeres vèrtebres, el programari permet tornar invisible la primera vèrtebra i importar una nova vèrtebra. Ara podem veure només el segon i el tercer os, i podem ajustar la posició del tercer os perquè encaixe perfectament amb el segon os. Fer-ho així elimina qualsevol biaix que les nocions preconcebudes pogueren introduir en la postura neutra de la columna vertebral. Aquestes nocions preconcebudes encara avui dia influeixen moltíssim en la manera com s’articulen els esquelets de sauròpodes en nombrosos museus, on moltes vegades s’ha de forçar la desarticulació de la columna vertebral en diversos punts per aconseguir una postura amb l’esquena arquejada, per exemple.
Sorprenentment, l’esquelet virtual resultant d’aplicar aquest protocol fou radicalment diferent de totes les reconstruccions anteriors d’aquest dinosaure, que mostraven Spinophorosaurus amb una columna vertebral horitzontal. No obstant això, la nova reconstrucció mostrava un coll i una esquena inclinats cap amunt amb una cua horitzontal (Vidal, Mocho, Aberasturi et al., 2020; Vidal, Mocho, Páramo et al., 2020). Tot i la seua elevació, el coll quedava alineat pràcticament amb les vèrtebres dorsals, sense canvis en l’angle d’elevació a la base del coll, com havien demostrat estudis anteriors (Stevens i Parrish, 2005). Però com és possible que Spinophorosaurus tinga el coll inclinat cap amunt? Hi hauria d’haver un punt on la columna vertebral s’enlairara, on estiguera la «clau de volta» dels sauròpodes. I trobem aquesta pedra angular a les vèrtebres del sacre, les vèrtebres fusionades amb els ossos del maluc.
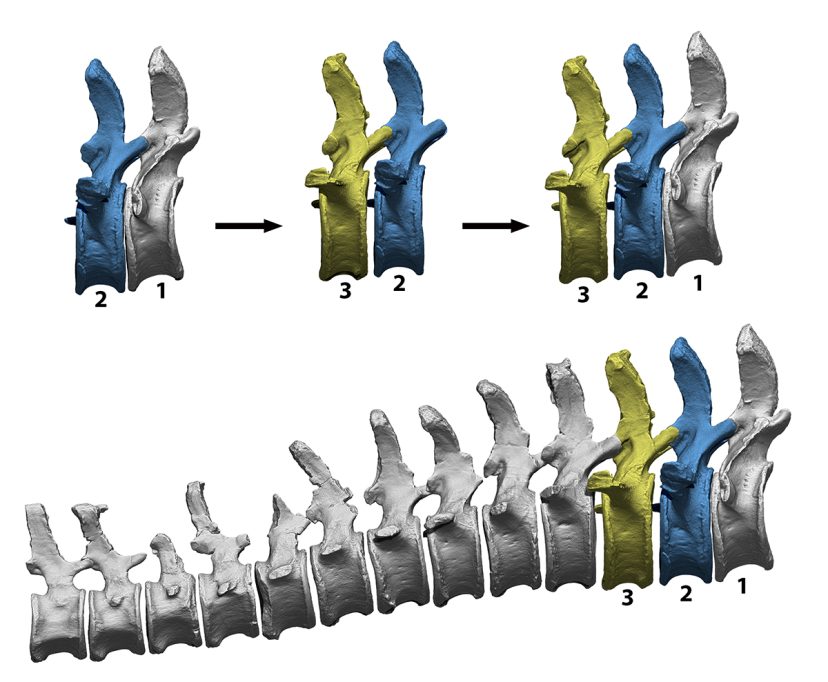
Procés d’articulació en postura neutra d’una seqüència de vèrtebres caudals virtuals del dinosaure sauròpode Camarasaurus, amb el qual evitar que nocions preconcebudes tinguen un impacte significatiu en la curvatura de la columna vertebral. Primerament, s’articula el primer parell de vèrtebres, de manera que el solapament de les superfícies d’articulació és màxim. Després s’oculta la primera vèrtebra caudal i s’hi afegeix la tercera, mentre sempre estan visibles una parella i s’articulen mitjançant el mateix criteri. La curvatura resultant és fruit del màxim contacte articular, cosa que minimitza possibles biaixos d’interpretació. / Daniel Vidal
Això va resultar sorprenent, ja que la majoria dels paleontòlegs esperaven que el punt clau fos on el coll s’uneix amb l’esquena o la base del coll (Stevens, 2013; Taylor et al., 2009). Tot i això, com va dir el paleontòleg Henry F. Osborn (1899), com a les aus, «la unió sacropèlvica és el centre de l’energia i del moviment», és l’àrea més reforçada de la columna vertebral; per tant, la que podria suportar modificacions sense comprometre’n la capacitat perquè l’animal caminara. El sacre de Spinophorosaurus té forma de falca o pedra angular: és més llarg a la part inferior i més curt a la meitat superior. En conseqüència, quan s’articula la cua, el tors i el coll s’eleven cap amunt, la qual cosa permetia que des d’aquesta postura neutra Spinophorosaurus situara el musell a cinc metres de terra, més del doble d’alçada que la seua espatlla.
Eren els sauròpodes com les girafes?
Per saber si aquestes tècniques realment són capaces de desentranyar dades precises de la paleobiologia dels dinosaures sauròpodes, l’aproximació més immediata és avaluar si és possible replicar-hi les capacitats biològiques de vertebrats actuals. La girafa és l’únic vertebrat actual que es pot utilitzar com a model per als colls dels sauròpodes: són animals quadrúpedes de grans dimensions que superen la tona de pes i amb un llarg coll, fruit de l’elongació de les vèrtebres cervicals i la «cervicalització» de la seua primera vèrtebra toràcica. Si s’apliquen sobre esquelets de girafes actuals les mateixes tècniques descrites per als sauròpodes, les reconstruccions esquelètiques obtingudes coincideixen perfectament amb les postures habitualment adoptades per les girafes en caminar (Vidal, Mocho, Páramo et al., 2020).
A la natura, les girafes s’alimenten habitualment amb el coll situat per sobre de les espatlles, però a vegades els mascles s’alimenten també, amb el coll completament vertical, dels brots més alts als quals puguen arribar i les femelles s’alimenten pasturant arran de terra. Tot aquest rang de posicions dels colls de les girafes es pot obtenir amb els seus esquelets virtuals sense necessitat de desarticular cap vèrtebra. A més, fins i tot la postura de màxima ventriflexió del coll de les girafes virtuals no permet arribar a terra sense flexionar els colzes o espatarrar les espatlles, exactament igual que passa amb les girafes vives. Es pot afirmar que tot el repertori de posicions del coll de les girafes es pot replicar virtualment amb els mateixos paràmetres utilitzats per als sauròpodes.
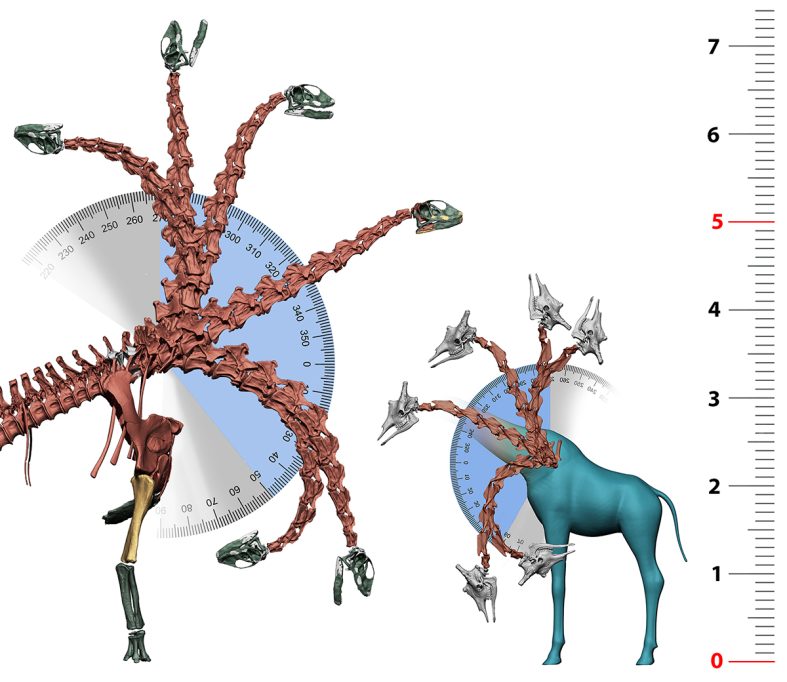
Què implica això per als dinosaures sauròpodes? Aquesta és una qüestió més complexa que afirmar que si un organisme extint podia adoptar físicament una postura, aquesta era adoptada habitualment i formava part del seu comportament. Els éssers humans som capaços de caminar sobre la punta dels dits dels peus quan ballem ballet i els elefants són capaços d’aguantar-se sobre les dues potes davanteres. Això no implica, però, que ballar ballet o aguantar-se amb les extremitats davanteres siguen comportaments habituals de cap d’aquestes dues espècies (Mallison, 2010; Sander et al., 2010). No obstant això, aquesta capacitat existeix. Les tècniques de paleontologia virtual permeten desvelar quines capacitats físiques tenien vertebrats extints i quines postures o comportaments eren físicament impossibles i, per tant, s’haurien de descartar a l’hora d’interpretar l’ecomorfologia d’aquests organismes (Mallison, 2010). En el cas de Spinophorosaurus nigerensis es pot afirmar que el seu esquelet li permetia adoptar posicions que li hagueren permès alimentar-se igual que les girafes actuals: des de copes d’arbres fins a 8 metres d’alçada fins a vegetació a nivell de terra. És un dels primers sauròpodes dels quals tenim evidència d’aquestes capacitats. Altres sauròpodes, com els diplodòcids Apatosaurus, Diplodocus o Amargasaurus, no tenien esquelets que els conferissen les mateixes capacitats de moviment i estaven més restringits a l’hora de situar el coll verticalment, cosa que evidencia que probablement ocuparien nínxols ecològics diferents (Christian et al., 2005; Stevens i Parrish, 1999, 2005). Per tant, els models virtuals sí que permeten reproduir les capacitats funcionals dels vertebrats actuals i, consegüentment, també inferir les dels vertebrats extints. Tot i això, no és possible saber si totes aquestes capacitats formaven part del seu comportament o, si era així, si eren capacitats usades habitualment o de forma esporàdica. Tanmateix, és possible utilitzar aquestes tècniques per refutar comportaments, sempre que aquests siguen incompatibles amb les capacitats d’un organisme, i la paleontologia virtual és capaç de mostrar-ho.
La còpula en els dinosaures sauròpodes
Un misteri difícil de conèixer és la interpretació correcta de com es reproduïen els dinosaures no aviaris. És segur que els dinosaures, fins i tot els sauròpodes més grans, podien fer la còpula, ja que tots van tenir ascendència i descendència, però la seua grandària gegantina ens fa preguntar-nos com era exactament. Per fer-se una idea de com podria ser aquesta còpula, el primer és determinar com la fan els parents vius més propers als dinosaures sauròpodes, és a dir, els cocodrils i les aus. La còpula en cocodrils és molt similar en totes les espècies actuals i és la mateixa que en molts rèptils: el mascle se situa al costat de la femella i col·loca la seua extremitat posterior sobre el maluc de la femella, una posició anomenada en anglès leg over back (literalment, ‘cama sobre l’esquena’). Aquesta posició facilita que el «penis» del mascle es puga introduir a la claveguera de la femella, ja que l’apèndix caudal dels rèptils sol dificultar la còpula (Isles, 2009). Les aus reduïren moltíssim el seu apèndix caudal durant la seua evolució, i en la major part d’aus actuals l’apèndix està format per unes poques vèrtebres amb mobilitat, de manera que no suposa un impediment per a la còpula. Per aquest motiu, les aus no fan la postura de leg over back com sí que fan els cocodrils. Les aus més primitives tenen un «penis» (de funció anàloga al dels mamífers, encara que d’origen evolutiu diferent), però els passeriformes i els gal·liformes tendeixen a reduir-lo o perdre’l del tot (Isles, 2009).
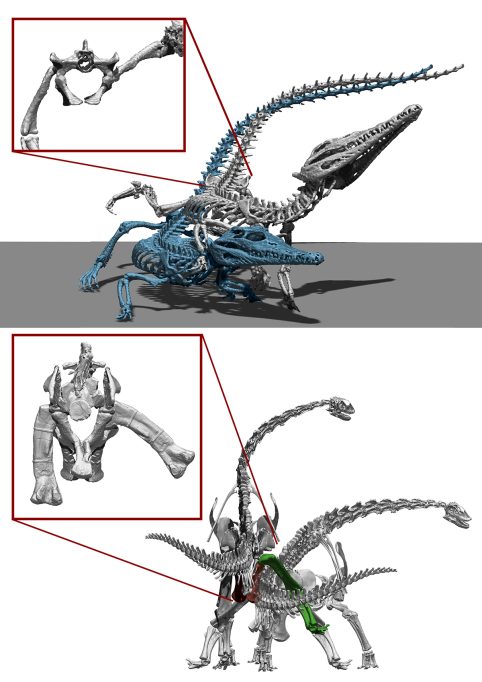
Els cocodrils actuals tenen la capacitat d’aixecar en abducció la seua extremitat posterior molt més enllà de l’horitzontal, cosa que els permet fer la còpula en la posició leg over back. Però els dinosaures sauròpodes amb prou feines podien aixecar en abducció l’extremitat posterior, ja que l’epífisi proximal del fèmur col·lidiria amb les parets de l’acetàbul i es produiria una parada osteològica. Això suposa que els sauròpodes eren incapaços d’una còpula típica d’una gran majoria de rèptils actuals. / Daniel Vidal
El fet que els cocodrils tinguen un pla corporal més similar al dels sauròpodes (tenen un apèndix caudal ben desenvolupat i són quadrúpedes) suggereix que potser aquests últims adoptaren també el leg over back com a estratègia per facilitar la còpula (Hallet i Wedel, 2016; Isles, 2009). Tanmateix, l’esquelet dels sauròpodes simplement no permetria aquesta posició. A diferència dels cocodrils, el fèmur dels quals té un cap arrodonit que permet molta mobilitat a l’acetàbul del maluc, els sauròpodes tenen fèmurs amb un cap en forma d’ela invertida. Això fa que qualsevol intent d’abducció aviat es veja limitat en col·lidir amb les parets de l’acetàbul, reduint molt el seu moviment en aquest pla. La capacitat limitada dels sauròpodes per elevar la seua extremitat posterior vol dir que no hi hauria prou espai entre el maluc i l’extremitat posterior per acomodar la femella, com sí que passa en els cocodrils, llevat que aquesta fora molt més petita que el mascle. Hi ha pocs estudis sobre el dimorfisme sexual en dinosaures sauròpodes, però la poca evidència existent indica que no hi havia un dimorfisme molt acusat en la mida entre ambdós sexes (Ikejiri, 2004). Per tant, es pot assegurar que els sauròpodes no podien adoptar la mateixa posició de còpula que els cocodrils i una gran majoria de rèptils actuals, i que les seues estratègies de còpula devien ser diferents. Altres possibilitats que hagueren pogut adoptar els sauròpodes són una munta similar a la que fan la majoria dels mamífers quadrúpedes, en què el mascle se situaria sobre les seues extremitats del darrere i s’aguantaria amb les davanteres sobre la femella, que podria situar-se dreta o recolzada. No hi ha cap impediment en el rang de moviment osteològic dels sauròpodes que impedira dur a terme aquestes posicions. Tanmateix, tampoc no hi ha cap impediment osteològic que pogueren fer una còpula d’esquena, de manera similar a com la fan molts artròpodes i alguns cànids. En definitiva, no podem saber com s’aparellaven els sauròpodes, ja que no podem refutar totes les possibilitats. El que sí que podem afirmar amb absoluta certesa és que no ho feien de la mateixa manera que els cocodrils actuals.
Conclusió: una eina addicional i complementària
Un colp vistos aquests exemples, és evident que les tecnologies virtuals no són una novetat que ha arribat per substituir la paleontologia i l’anatomia comparades tradicionals per haver quedat obsoletes, sinó que són una eina addicional i complementària per poder avaluar científicament algunes hipòtesis que abans resultaven molt difícils o fins i tot impossibles de ser testades. Hem vist com la paleontologia virtual mostra que la hipòtesi tradicional dels sauròpodes com a anàlegs de les girafes era totalment matisable, i que alguns sauròpodes tenien capacitats per alimentar-se igual que les girafes, mentre que altres tenien capacitats de moviment més limitades en els seus colls, la qual cosa els condicionava a alimentar-se de vegetació de baix port o arran de terra. Això suggereix un escenari més complex als considerats prèviament, on no es tracta de si tots els sauròpodes podien o no podien alimentar-se com les girafes, sinó de quines podien i quines no ho podien fer.
De la mateixa manera, fins fa poc, com s’aparellaven els sauròpodes era una qüestió propera a l’especulació. Tot i això, gràcies a la paleontologia virtual s’ha pogut refutar que els sauròpodes s’aparellaren com ho fan els cocodrils actuals. La paleontologia virtual no ha deixat de requerir observacions en animals actuals vius o de comptar amb exemplars fòssils excepcionalment complets per poder avaluar aquestes hipòtesis de forma contundent. Ha suposat alhora una revolució en com estudiem els fòssils de dinosaures, però també un recordatori de la importància de la confluència de múltiples línies d’evidència com a millor manera d’avançar el coneixement científic.
Bakker, R. T. (1971). Ecology of the brontosaurs. Nature, 229, 172–174. https://doi.org/10.1038/229172a0
Dzemski, G., & Christian, A. (2007). Flexibility along the neck of the ostrich (Struthio camelus) and consequences for the reconstruction of dinosaurs with extreme neck length. Journal of Morphology, 268, 701–714. https://doi.org/10.1002/jmor.10542
Hallett, M., & Wedel, M. J. (2016). The sauropod dinosaurs. Johns Hopkins University Press.
Ikejiri, T. (2004). Anatomy of Camarasaurus lentus (Dinosauria: Sauropoda) from the Morrison formation (Late Jurassic), Thermopolis, Central Wyoming, with determination and interpretation of ontogenetic, sexual dimorphic and individual variation in the genus. Fort Hays State University.
Isles, T. E. (2009). The socio-sexual behaviour of extant archosaurs: Implications for understanding dinosaur behaviour. Historical Biology, 21(3–4), 139–214. https://doi.org/10.1080/08912960903450505
Mallison, H. (2010). CAD assessment of the posture and range of motion of Kentrosaurus aethiopicus Hennig 1915. Swiss Journal of Geosciences, 103(2), 211–233. https://doi.org/10.1007/s00015-010-0024-2
Mallison, H., & Wings, O. (2014). Photogrammetry in paleontology–A practical guide. Journal of Paleontological Techniques, 12, 1–31.
Osborn, H. F. (1899). A skeleton of Diplodocus, recently mounted in the American Museum. Science, 10(259), 870–874. https://doi.org/10.1126/science.10.259.870
Remes, K., Ortega, F., Fierro, I., Joger, U., Kosma, R., Ferrer, J. M. M., Ide, O. A., & Maga, A. (2009). A new basal sauropod dinosaur from the middle Jurassic of Niger and the early evolution of Sauropoda. PLOS ONE, 4(9), e6924. https://doi.org/10.1371/journal.pone.0006924
Sander, P. M., Christian, A., Clauss, M., Fechner, R., Gee, C. T., Griebler, E.-M., Gunga, H.-C., Hummel, J., Mallison, H., Perry, S. F., Preuschoft, H., Rauhut, O. W. M., Remes, K., Tutken, T., Wings, O., & Witzel, U. (2010). Biology of the sauropod dinosaurs: The evolution of gigantism. Biological Reviews, 86, 117–155. https://doi.org/10.1111/j.1469-185X.2010.00137.x
Sereno, P. C., Wilson, J. A., Witmer, L. M., Whitlock, J. A., Maga, A., Ide, O., & Rowe, T. A. (2007). Structural extremes in a cretaceous dinosaur. PLOS ONE, 2(11), 1–9. https://doi.org/10.1371/journal.pone.0001230
Stevens, K. A. (2013). The articulation of sauropod necks: Methodology and mythology. PLOS ONE, 8(10), 1–27. https://doi.org/10.1371/journal.pone.0078572
Stevens, K. A., & Parrish, M. J. (1999). Neck posture and feeding habits of two Jurassic sauropod dinosaurs. Science, 284, 798–800. https://doi.org/10.1126/science.284.5415.798
Stevens, K. A., & Parrish, M. J. (2005). Neck posture, dentition, and feeding strategies in Jurassic sauropod dinosaurs. En V. Tidwell & K. Carpenter (Eds.), Thunder-lizards: The sauropodomorph dinosaurs (p. 212–232). Indiana University Press.
Sutton, M. D., Rahman, I. A., & Garwood, R. J. (2014). Techniques for virtual palaeontology. Wiley. https://doi.org/10.1002/9781118591192
Taylor, M. P. (2010). Sauropod dinosaur research: A historical review. Geological Society London Special Publications, 343(1), 361–386. https://doi.org/10.1144/SP343.22
Taylor, M. P., Wedel, M. J., & Naish, D. (2009). Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica, 54(2), 213–220. https://doi.org/10.4202/app.2009.0007
Vidal, D., Mocho, P., Aberasturi, A., Sanz, J. L., & Ortega, F. (2020). High browsing skeletal adaptations in Spinophorosaurus reveal an evolutionary innovation in sauropod dinosaurs. Scientific Reports, 10(1), 6638. https://doi.org/10.1038/s41598-020-63439-0
Vidal, D., Mocho, P., Páramo, A., Sanz, J. L., & Ortega, F. (2020). Ontogenetic similarities between giraffe and sauropod neck osteological mobility. PLOS ONE, 15(1). https://doi.org/10.1371/journal.pone.0227537




