

Sexe i disseny en el nostres parents evolutius
La percepció de la bellesa en la natura
Des d’un enfocament evolutiu de la bellesa, en aquest text analitzem l’expressió i la percepció de la bellesa sexual en el regne animal. Els cervells dels animals perceben la bellesa i detecten les característiques de l’entorn que resulten més rellevants per a la seua supervivència. Al llarg de l’evolució, els animals amb reproducció sexual han explotat aquest procés de detecció per a maximitzar el seu atractiu per a l’altre sexe. Habitualment, això ha comportat l’aparició de comportaments i trets de festeig extrems. Aquests són trets de bellesa sexual. Combinant principis neurocientífics i neuroestètics moderns amb altres provinents de la biologia evolutiva, tractem d’entendre les bases biològiques i l’evolució de la bellesa en tots els animals, inclosos nosaltres mateixos.
Paraules clau: bellesa, neuroestètica, fluïdesa perceptiva, biaix sensorial, selecció sexual.
Diuen que la bellesa és als ulls de qui mira, però, més enllà de l’aforisme, és difícil donar-ne una definició clara. De fet, històricament, tant a filòsofs com a científics els ha costat descriure la bellesa, i avui dia continuem buscant-ne el significat. A causa d’una manca d’una definició sòlida o consensuada, podem pensar que no hi ha hagut grans progressos en aquest sentit, però això no és del tot cert. Els primers filòsofs grecs van suggerir que la bellesa era una qualitat objectiva, una propietat inherent de les coses belles. Si un objecte s’ordenava i s’organitzava perfectament al voltant de la seua funció, era bell per definició, independentment de qui el percebera. Avui dia, pocs científics estan d’acord amb aquesta idea. La majoria considera que la bellesa no és objectiva, sinó subjectiva, una propietat emergent de la nostra ment en interaccionar amb l’entorn. Responem a objectes i sons del món real, sí, però el procés d’experimentar la bellesa es produeix en la nostra ment. Quan parlem de bellesa, parlem de cervells. De vegades, la bellesa pot estar als ulls de qui la mira, però està en tot moment en el seu cervell.
La disciplina moderna de la neuroestètica es va desenvolupar entorn d’aquest paper central del cervell. La neuroestètica va començar com l’estudi de les respostes humanes davant l’art, principalment el visual, però la disciplina ha crescut i ja inclou no solament diferents formes artístiques, com la música i la poesia, sinó també estímuls més rellevants en termes biològics, com els paisatges i els rostres. La neuroestètica s’ha expandit gràcies als avanços en neurociència i ha incorporat mètriques d’activitat neuronal cada vegada més sofisticades, com l’electroencefalografia (EEG), la tomografia per emissió de positrons (PET) i la imatge per ressonància magnètica funcional (IRMf). La neuroestètica és, per tant, un dels camps d’estudi interdisciplinaris per excel·lència, i connecta personal acadèmic de les humanitats, la neurociència, la psicologia, la informàtica i la biologia evolutiva. Aquí considerem la qüestió de la bellesa des de la perspectiva d’aquesta última.
Com a éssers humans, trobem bellesa al nostre voltant. Com a biòlegs evolutius, ens preguntem on es va originar aquest «gust per la bellesa», i com a ecòlegs del comportament, ens preguntem fins a quin punt aquest gust es manifesta en altres animals, i quina mena de coses consideren aquests que són belles. Per als éssers humans, l’objecte bell amb més importància biològica és, possiblement, el rostre d’un altre ésser humà, especialment aquells pels quals sentim una atracció sexual. En conseqüència, un candidat probable a la bellesa en el cas dels animals són els trets que utilitzen per atreure membres del sexe oposat (Prum, 2017; Ryan, 2018). Aquests trets, coneguts com a caràcters sexuals secundaris, són omnipresents en la natura i, en alguns animals, com el quetzal esplèndid (Pharomachrus mocinno), s’han dut a l’extrem (Figura 1).

Figura 1. Mascle de quetzal esplèndid (Pharomachrus mocinno). Els mas- cles d’aquesta espècie es caracteritzen pels colors cridaners, ornaments que utilitzen per a competir per l’atenció de les femelles./ Harleybroker
Selecció sexual
Els caràcters sexuals secundaris s’han associat amb la ciència de la bellesa com a mínim des de Charles Darwin, qui va afirmar que «els poders de percepció de l’home i dels animals inferiors estan constituïts de tal forma que els colors lluents i certes formes, així com els sons harmoniosos i rítmics, es consideren bells; però de per què això és així no en sabem més […]». 1 Si substituïm home per persones i inferiors per altres, la pregunta continua vigent quan investiguem l’origen i la base biològica de la bellesa en tot el regne animal. De fet, el seu enfocament quant al plaer emfatitzava adequadament el paper del cervell com l’òrgan sexual més important, i constitueix la base de la seua teoria de la selecció sexual (Darwin, 1871). Malgrat que l’Anglaterra victoriana es resistia a admetre-ho (Richards, 2017) probablement aquesta teoria era correcta des del primer moment, ja que se centrava en el poder de la ment per a generar bellesa en la natura.
La selecció sexual és responsable de generar gran part de la bellesa del món animal. Els trets que evolucionen per selecció natural milloren la capacitat d’un animal per a aconseguir parella; alguns d’aquests són armaments i d’altres són ornaments. Els armaments faciliten el combat per a obtenir parella, mentre que els ornaments evolucionaren per a atraure els membres del sexe oposat i són, per tant, trets de bellesa sexual (Rosenthal, 2017). Darwin es va centrar en els sistemes d’aparellament més típics: mascles –els «festejadors»– que utilitzen ornaments per a competir per l’atenció de les femelles –les «selectores»–, que decideixen amb qui s’aparellen (Darwin, 1871). Però des de temps de Darwin, s’han descobert variacions substancials en aquest procés, incloent-hi trets que funcionen alhora com a ornaments i com a armaments, espècies en què les femelles són les festejadores i els mascles els selectors, i molts sistemes d’aparellament –especialment en els éssers humans– en què ambdós sexes seleccionen parella (Rosenthal i Ryan, 2022).
Deixant-ne de banda la bellesa, la producció i el manteniment dels ornaments sexuals solen ser costosos i, en molts casos, atrauen no només les parelles, sinó també els depredadors. Darwin va suggerir que aquests trets podien continuar evolucionant si el benefici de l’atracció superava el cost d’una menor supervivència (Darwin, 1871). Gran part de l’ecologia del comportament s’ha preguntat quins beneficis aconsegueixen els selectors per triar trets bells. Però aquí ens interessa què és el que fa bells aquests trets en primer lloc. Darwin ens va donar algunes pistes al respecte: «Quan un animal mascle emet sons amb la finalitat d’agradar a les femelles, naturalment utilitzarà aquells que siguen dolços a l’oïda de l’espècie» (Darwin, 1872).2 Donat que la bellesa és al cervell de qui l’observa, moltes investigacions recents sobre l’elecció de parella han explorat l’arquitectura neuronal, perceptiva i cognitiva que subjau al gust per la bellesa.
La base neuronal i social de la bellesa
Encara que el cervell siga l’òrgan sexual més important, aquest no és l’únic tema en què se centra. L’arquitectura neuronal d’un cervell no es constitueix únicament entorn del sexe, sinó que es veu influenciada pel nínxol ecològic multidimensional en què ha evolucionat. La Terra es compon d’un nombre incommensurable de nínxols ecològics diferents, als quals els cervells poden adaptar-se –fins i tot en el mateix nínxol, un únic problema es pot solucionar de formes distintes– i això dona lloc a una diversitat proporcional d’arquitectures neuronals. Una presa amenaçada durant el dia, per exemple, pot dedicar una quantitat significativa de recursos mentals a la visió d’espectre complet per a detectar millor els depredadors. Alternativament, podria invertir més recursos en habilitats propioceptives i motrius amb què fugir-ne de forma més eficaç. Aquestes especialitzacions neuronals creen el que Von Uexküll anomenava l’umwelt de l’animal: el «món interior» a través del qual percep i interactua amb el seu entorn (Von Uexküll, 2014). La diversitat de nínxols i adaptacions han donat lloc a una sorprenent varietat en els mons interiors dels animals, i aquestes adaptacions neuroecològiques creen biaixos –en sensacions, percepcions, cognicions o decisions– que dicten allò que un animal considera atractiu o bell.
La percepció d’alguna cosa com a bella es forma en el cervell, però abans l’essència de la bellesa ha de passar per un procés de transducció a respostes neuronals per part dels òrgans sensorials del subjecte. Aquests òrgans (orelles, ulls, narius i altres) són els portals a través dels quals els sons, les imatges, les olors i altres sensacions arriben al cervell. Però no per això els estímuls tenen via lliure. Els òrgans sensorials actuen com a guardians i només permeten passar una part d’aquests estímuls. Tots els òrgans sensorials estan calibrats d’una determinada manera i són més sensibles a uns estímuls que a altres. Nosaltres, per exemple, no percebem els colors ultraviolats del plomatge de les aus, ni sentim les crides ultrasòniques d’ecolocalització dels ratpenats, i només podem detectar part del conjunt d’olors que perceben els gossos. Naturalment, els animals desenvolupen senyals de festeig que els seus propis sistemes sensorials poden percebre, en un procés que coneixem com explotació sensorial (Ryan, 2018).

Figura 2. Granota túngara (Engystomops pustulosus) mascle fent una crida. Els animals desenvolupen estratègies de festeig en funció dels estímuls que poden percebre. La granota túngara i el seu parent més proper són els únics que utilitzen la papil·la basilar per a comunicar-se, i el to de la seua crida va evolucionar per a coincidir amb els biaixos previs de la papil·la basilar. Aquesta estimulació sensorial extra fa que les seues crides resulten més atractives sexualment a les femelles./ Foto: Ryan Taylor
Per exemple, les granotes tenen dos òrgans auditius diferents per a processar el so, la papil·la amfíbia i la papil·la basilar, i ambdós òrgans perceben tons distints. La granota túngara Engystomops (= Physalaemus) pustulosus (Figura 2) i altres espècies properes fan unes crides similars a un gemec que estimulen la papil·la amfíbia, però només la granota túngara i la seua espècie més propera (Engystomops petersi) hi afegeixen síl·labes (anomenades chucks), que coincideixen amb els sons que detecta la papil·la basilar. Aquests sons multipliquen per cinc l’atractiu sexual de la crida! Encara que només aquestes dues espècies utilitzen la papil·la basilar per a comunicar-se, totes les espècies properes tenen papil·les basilars equipades per a detectar aquests sons. Per tant, el to dels chucks va evolucionar per a ajustar-se al biaix previ que suposava la papil·la basilar (Wilczynski et al., 2001). Aquesta estimulació sensorial extra fa que les seues crides resulten més atractives sexualment a les femelles.
Existeixen proves de molts patrons anàlegs en la comunicació visual (Cummings i Endler, 2018). En el llac Victòria, per exemple, el color i la il·luminació de l’entorn varien amb la profunditat. Els peixos cíclids que hi viuen a diferents profunditats tenen, per tant, sensibilitats distintes, perquè els colors als quals estan més habituats els seus ulls són els de l’entorn nadiu. I, en conseqüència, els colors que mostren en el festeig han evolucionat per a ajustar-se a aquests ulls. En un exemple estrany que té a veure amb les olors, els mascles de les abelles de les orquídies intenten aparellar-se amb orquídies que imiten l’olor de les abelles femella per a atreure’ls a les seues flors i que actuen com a pol·linitzadors inadvertidament (vegeu Ryan, 2018).

Figura 3. Una mostra del ventall de coloració dels guppys mascle de Trinitat (Poecilia reticulata). Aquest peix és conegut no només per la varietat de colors dels mascles, sinó també per la varietat de patrons en què es disposen aquests colors. Pot ser que els guppys aprofiten un biaix perceptiu en favor de l’alt contrast, que es pot aconseguir gràcies a la varietat de patrons.
Una vegada que aquests estímuls (o, més concretament, les respostes neuronals que hi susciten) arriben al cervell, se sotmeten a un processament superior que dona lloc a les percepcions de bellesa de l’animal. I quan parlem de la percepció de la bellesa, el peix guppy (Poecilia reticulata) n’és sens dubte l’estrella (Figura 3). Aquests acolorits peixets són els conillets d’Índies de la investigació en selecció sexual. Són coneguts no només per la varietat de colors brillants dels mascles, sinó també pel ventall de patrons en què es configuren. Com en l’art, els colors no en són l’únic element rellevant; també ho és la forma en què aquests es disposen, i els patrons es processen de forma unitària com perceptes. Una de les explicacions de l’ampli espectre de patrons de color dels guppys és la gran sensibilitat al color dels ulls d’aquests peixos. És possible, per exemple, que individus distints perceben els colors de forma diferent (Houde, 1997). O pot ser que els guppys aprofiten un biaix perceptiu en favor de l’alt contrast, que es pot aconseguir gràcies a la varietat de patrons. Sibeaux et al. (2019) van descobrir que les femelles senten atracció pels mascles amb cossos amb més variació contrastiva. És possible que aquesta variació evite l’adaptació sensorial, una espècie d’avorriment neurològic, quan les femelles exploren els cossos dels mascles.
Es diu que Marilyn Monroe va defensar que «és millor ser absolutament ridícul que absolutament avorrit». L’avorriment és la sentència de mort de la bellesa sexual, i els patrons també són importants en el context de l’habituació, la versió conductual de l’avorriment. Això va quedar raonablement clar en un experiment simple però elegant amb guppys (amb qui, si no). Els investigadors van criar dos llinatges de guppys en el laboratori. Els mascles d’ambdues línies no presentaven diferències notables en el tipus o quantitat de colors que expressaven, però sí en la seua disposició, és a dir, en els patrons. Es va observar una disminució en la resposta sexual de les femelles quan se’ls presentaven mascles de la seua mateixa línia, i la resposta es recuperava davant els mascles de l’altra línia, que mostraven un patró nou (Daniel et al., 2019).
També es pensa que la lluita contra l’habituació pot explicar l’evolució del cant de les aus (Hartshorne, 1973). Quan s’exposa a nous tons de cant als diamants zebrats de Timor, tant la seua activitat cerebral com el seu comportament sexual augmenten (Dong i Clayton, 2009). Com ocorre amb la relació entre colors i la seua disposició, els patrons de so són fonamentals per a la bellesa del cant de l’au, i això és cert tant per a les aus com per a nosaltres. Bilger et al. (2021) van demostrar que els patrons de so naturals en el cant de les aus ens resulten més bells que les mateixes notes disposades en un ordre aleatori. Aquests estudis suggereixen que les aus i els humans comparteixen l’atracció per patrons musicals d’ordre superior. Darwin ho va predir, per descomptat, especialment en relació amb el plomatge i el cant. Pensava que era possible que les aus i els humans compartirem les mateixes preferències estètiques: «En resum, els ocells semblen ser els més estètics de tots els animals a excepció de l’home, i tenen per la bellesa un gust semblant al nostre»3 (Darwin, 1871). Aquestes influències de l’habituació subratllen més encara que la bellesa no és una qualitat inherent dels objectes, sinó de la forma en què es perceben.
Ara ja està clar que les percepcions de bellesa sexual poden ser mal·leables; per exemple, el comportament d’altres pot canviar les nostres percepcions. En un experiment que ja s’ha convertit en un clàssic, Dugatkin i Godin (1992) van demostrar el que ara coneixem com a «còpia de l’elecció de parella». En un experiment controlat en el laboratori, es permet a una guppy femella triar entre dos mascles. En tria un, el mascle preferit. A continuació, observa el mascle no triat festejant una femella model. Es retira la femella model i aleshores la femella inicial canvia la seua preferència en favor del mascle que abans no li interessava. Ara sabem que la còpia d’elecció de parella està molt estesa en el regne animal, també entre nosaltres.
Per exemple, l’avaluació d’un home fotografiat amb una dona atractiva és superior a la del mateix subjecte per si mateix. Però, per què? Recentment, Street et al. (2018) van demanar als participants d’un estudi que puntuaren l’atractiu d’una fotografia. Van rebre informació sobre com havien valorat la mateixa foto altres participants, i després se’ls va donar oportunitat de canviar la seua puntuació. Per regla general, els participants van canviar la seua puntuació en un 13 % cap a l’opinió majoritària. Els investigadors van dur a terme el mateix estudi perquè els participants puntuaren l’atractiu de mans i d’obres d’art. Sorprenentment, el canvi del 13 % cap a l’opinió majoritària es va produir en ambdós experiments. La conclusió és que sí, la còpia d’elecció de parella ocorre també en humans, però pot ser que siga una manifestació general de la facilitació social (Street et al., 2018). En conjunt, aquests estudis mostren que els cervells animals estan esbiaixats, tant pel que fa a sensacions com a percepcions i facilitació social. El nostre cervell ha sigut modelat per la nostra història evolutiva, que pot ser aprofitada per possibles parelles per a augmentar la seua bellesa percebuda.
Gust i desig: la bellesa i el sistema de recompensa
Una nova i emocionant línia d’investigació en l’estudi de la bellesa és la relacionada amb el paper del sistema de recompensa del cervell. Els nostres sistemes de recompensa són regions del cervell raonablement ben definides responsables del fet que ens «agrade» alguna cosa (la recompensa) i que la «desitgem» i tractem d’aconseguir-la. Berridge i Robinson van demostrar clarament el funcionament d’aquests mecanismes psicològics diferenciats, el gust i el desig per alguna cosa, en un estudi amb rates de laboratori, l’espècie model de la psicologia comparativa (Berridge i Robinson, 1998). A les rates els agrada el sucre. Però si estan mancades de dopamina (si les neurones dopaminèrgiques són destruïdes mitjançant la inoculació d’una neurotoxina), ja no en desitgen; és a dir, deixen de treballar per a obtenir-ne, malgrat que, quan un investigador els en dona, el seu gaudi és evident. El marc gust-desig pot ser important per a la nostra comprensió de la bellesa, ja que, possiblement, l’únic punt de consens sobre la bellesa és que aquesta implica alguna mena de plaer. Per tant, la capacitat de mesurar el plaer com un procés fisiològic en si mateix és fonamental per a comprendre les respostes a estímuls sexuals. És evident que els animals clarament «desitgen» el sexe; probablement, també els «agrada». Una pregunta que paga la pena fer és si els caràcters sexuals secundaris, responsables de tanta bellesa en la natura, provoquen plaer («agraden») als animals a què van destinats.
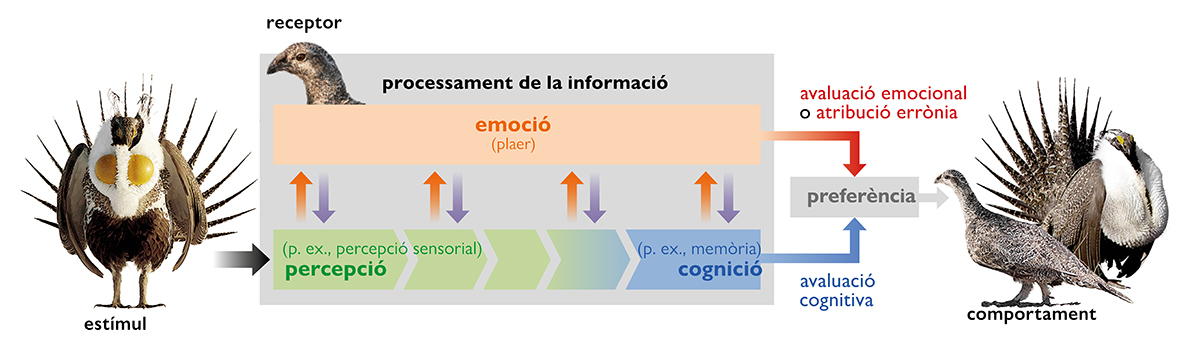
Moltes proves indiquen que els caràcters sexuals secundaris activen els circuits de recompensa dels animals (Lynch i Ryan, 2020). Per exemple, els cants d’aparellament dels mascles activen la via mesolímbica de recompensa en espècies tan allunyades com el sit gorjablanc (Maney, 2013) i la granota túngara (Hoke et al., 2010). Aquests i altres estudis deixen clar que els caràcters sexuals secundaris activen el desig, o n’augmenten els incentius. També hi ha proves que els caràcters sexuals secundaris poden provocar plaer («agradar») independentment de la seua habilitat per a provocar «desig». Utilitzant dues respostes conductuals diferents, Dai et al. (2010) van descobrir que la gent jutja per separat el «gust» i el «desig» en relació amb un rostre humà. Per tant, un interessant camí a seguir és desenvolupar estudis com aquests en animals no humans. Aquesta mena d’estudis ens poden indicar si els caràcters sexuals secundaris activen el gust a més del desig, cosa que seria d’esperar si els animals troben aquests trets tan bells com ho fem nosaltres.
Les proves teòriques suggereixen que així és. Els psicòlegs experimentals fan servir el terme fluïdesa perceptiva per a descriure les emocions positives generades per la informació que el cervell processa sense dificultat. Per exemple, a la gent li solen agradar les imatges simètriques o amb línies o superfícies contínues, que són fàcils de codificar per al cervell. Igualment, els investigadors en estètica empírica han demostrat que les imatges que comparteixen estadístiques especials amb les escenes naturals (els patrons i les formes que trobem en la natura) són més fàcils de processar pel cervell i reben valoracions més positives (agraden més). Fins i tot els caràcters de les nostres llengües escrites imiten les formes de les escenes naturals (Changizi et al., 2006). Els nostres cervells han evolucionat en la natura, i estan calibrats per a detectar-hi els patrons més importants per a la nostra supervivència i reproducció; no és estrany que puguem processar patrons naturals amb més facilitat. I quan el processament de la informació és senzill, experimentem plaer: ens «agrada». Com a éssers humans, sentim literalment atracció per imatges que «ens entren bé» pels ulls.

Figura 5. Variació de colors i patrons en algunes espècies de perca nord-americanes. Una de les qüestions pendents en relació amb la bellesa dels ornaments animals és fins a quin punt el biaix de processament pot explicar-ne l’atractiu. En els humans, les proves pareixen indicar que els rostres fàcils de processar es consideren més atractius./ Samuel Hulse
És possible que altres animals a banda dels humans compartisquen aquest biaix pels patrons naturals? Podria això explicar la bellesa dels caràcters sexuals secundaris? Renoult i Mendelson (2019) pensen que així és, com a mínim en part. Descriuen la preferència per patrons fàcilment processables com un «biaix de processament», que s’afig a la llista anterior de biaixos cap a determinats colors, sons i patrons. El biaix de processament podria ser particularment rellevant per a la percepció de la bellesa, ja que identifica explícitament el valor del plaer en el processament eficient de la informació (Figura 4). Aquest biaix podria ser aprofitat pels festejadors per a augmentar el seu atractiu, per exemple, quan els caràcters sexuals secundaris imiten les estadístiques espacials de les escenes naturals. Entre les proves que donen suport a aquesta hipòtesi, existeixen estudis sobre rostres humans que mostren que els rostres més senzills de processar es consideren més atractius (Holzleitner et al., 2019; Renoult et al., 2016), i un estudi amb peixos de colors (Figura 5) que va mostrar que els bells patrons de reproducció dels mascles imiten les estadístiques espacials de l’hàbitat en què van evolucionar (Hulse et al., 2020). Una altra línia d’investigació apassionant és fins a quin punt el biaix de processament pot explicar la bellesa dels ornaments animals, com pareix ser el cas amb els dissenys humans.
Observacions finals: comprendre el significat de la bellesa
Pel que fa a la bellesa, tenim molt a aprendre dels nostres parents evolutius. En la vida, com en l’art, les coses belles estan dissenyades per a produir plaer. Tant en el cas del quetzal al llarg de la seua evolució o de Georgia O’Keeffe en un calorós dia d’estiu, els cervells dels animals són alhora els creadors i els receptors de la bellesa en la natura. Per descomptat, part de la bellesa natural ens resulta invisible als humans, ja que ha evolucionat tenint en compte altres beneficiaris. La diversitat ecològica genera diversitat neurològica, que al seu torn genera varietat en la bellesa especialitzada en el nínxol en què va evolucionar. Però tenim la sort de continuar compartint molts dels biaixos perceptius i cognitius dels nostres parents evolutius, i la seua bellesa és també la nostra. Potser no hem arribat tan lluny com ens agradaria en la nostra cerca del significat de la bellesa, però la integració de nous coneixements del camp de la neuroestètica amb els principis establerts de la biologia evolutiva és una via prometedora.
Notes
1.Traducció de Josep Egozcue (L’origen de l’home i sobre la selecció en relació amb el sexe, vol. I, 1984, p. 280. Edicions Científiques Catalanes). (Tornar al text)
2.Traducció de Xavier Bellés (L’expressió de les emocions en l’home i en els animals, 2017, p. 61–62. Institut d’Estudis Catalans). (Tornar al text)
3.Traducció de Josep Egozcue (L’origen de l’home i sobre la selecció en relació amb el sexe, vol. I, 1984, p. 38. Edicions Científiques Catalanes). (Tornar al text)
Referències
Berridge, K. C., & Robinson, T. E. (1998). What is the role of dopamine in reward: Hedonic impact, reward learning, or incentive salience? Brain Research Reviews, 28(3), 309–369. https://doi.org/10.1016/S0165-0173(98)00019-8
Bilger, H. T., Vertosick, E., Vickers, A., Kaczmarek, K., & Prum, R. O. (2021). Higher-order musical temporal structure in bird song. Frontiers in Psychology, 12, 1–11. https://doi.org/10.3389/fpsyg.2021.629456
Changizi, M. A., Zhang, Q., Ye, H., & Shimojo, S. (2006). The structures of letters and symbols throughout human history are selected to match those found in objects in natural scenes. The American Naturalist, 167(5), E117–E139. https://doi.org/10.1086/502806
Cummings, M. E., & Endler, J. A. (2018). 25 years of sensory drive: The evidence and its watery bias. Current Zoology, 64(4), 471–484. https://doi.org/10.1093/cz/zoy043
Dai, X., Brendl, C. M., & Ariely, D. (2010). Wanting, liking, and preference construction. Emotion, 10(3), 324–334. https://doi.org/10.1037/a0017987
Daniel, M. J., Koffinas, L., & Hughes, K. A. (2019). Habituation underpins preference for mates with novel phenotypes in the guppy. Proceedings of the Royal Society B: Biological Sciences, 286, 20190435. https://doi.org/10.1098/rspb.2019.0435
Darwin, C. (1871). The descent of man and selection in relation to sex. John Murray.
Darwin, C. (1872). The expression of emotions in man and animals. John Murray.
Dong, S., & Clayton, D. F. (2009). Habituation in songbirds. Neurobiology of Learning and Memory, 92(2), 183–188. https://doi.org/10.1016/j.nlm.2008.09.009
Dugatkin, L. A., & Godin, J.-G. J. (1992). Reversal of female mate choice by copying in the guppy (Poecilia reticulata). Proceedings of the Royal Society B: Biological Sciences, 249, 179–184. https://doi.org/10.1098/rspb.1992.0101
Hartshorne, C. (1973). Born to sing. Indiana University Press.
Hoke, K. L., Ryan, M. J., & Wilczynski, W. (2010). Sexually dimorphic sensory gating drives behavioral differences in tungara frogs. Journal of Experimental Biology, 213(20), 3463–3472. https://doi.org/10.1242/jeb.043992
Holzleitner, I. J., Lee, A. J., Hahn, A. C., Kandrik, M., Bovet, J., Renoult, J. P., Simmons, D., Garrod, O., DeBruine, L. M., & Jones, B. C. (2019). Comparing theory-driven and data-driven attractiveness models using images of real women’s faces. Journal of Experimental Psychology. Human Perception and Performance, 45(12), 1589–1595. https://doi.org/10.1037/xhp0000685
Houde, A. E. (1997). Sex, color, and mate choice in guppies. Princeton University Press.
Hulse, S. V., Renoult, J. P., & Mendelson, T. C. (2020). Sexual signaling pattern correlates with habitat pattern in visually ornamented fishes. Nature Communications, 11(1), 1–8. https://doi.org/10.1038/s41467-020-16389-0
Lynch, K. S., & Ryan, M. J. (2020). Understanding the role of incentive salience in sexual decision-making. Integrative & Comparative Biology, 60(3), 712–721. https://doi.org/10.1093/icb/icaa054
Maney, D. L. (2013). The incentive salience of courtship vocalizations: Hormone-mediated ‘wanting’ in the auditory system. Hearing Research, 305(1), 19–30. https://doi.org/10.1016/j.heares.2013.04.011
Prum, R. O. (2017). The evolution of beauty: How Darwin’s forgotten theory of mate choice shapes the animal world-and us. Doubleday.
Renoult, J. P., Bovet, J., & Raymond, M. (2016). Beauty is in the efficient coding of the beholder. Royal Society Open Science, 3(3), 160027. https://doi.org/10.1098/rsos.160027
Renoult, J. P., & Mendelson, T. C. (2019). Processing bias: Extending sensory drive to include efficacy and efficiency in information processing. Proceedings of the Royal Society B: Biological Sciences, 286(1900), 20190165. https://doi.org/10.1098/rspb.2019.0165
Richards, E. (2017). Darwin and the making of sexual selection. University of Chicago Press.
Rosenthal, G. G. (2017). Mate choice: The evolution of sexual decision making from microbes to humans. Princeton University Press.
Rosenthal, G. G., & Ryan, M. J. (2022). Sexual selection and the ascent of women: Mate choice research since Darwin. Science, 375, eabi6308. https://doi.org/10.1126/science.abi6308
Ryan, M. J. (2018). A taste for the beautiful. Princeton University Press.
Sibeaux, A., Cole, G. L., & Endler, J. A. (2019). The relative importance of local and global visual contrast in mate choice. Animal Behaviour, 154, 143–159. https://doi.org/10.1016/j.anbehav.2019.06.020
Street, S. E., Morgan, T. J. H., Thornton, A., Brown, G. R., Laland, K. N., & Cross, C. P. (2018). Human mate-choice copying is domain-general social learning. Scientific Reports, 8(1), 1715. https://doi.org/10.1038/s41598-018-19770-8
Von Uexküll, J. (2014). Umwelt und Innenwelt der Tiere. Springer-Verlag.
Wilczynski, W., Rand, A. S., & Ryan, M. J. (2001). Evolution of calls and auditory tuning in the Physalaemus pustulosus species group. Brain, Behavior and Evolution, 58(3), 137–151. https://doi.org/10.1159/000047268




