

El mapa de la biodiversitat
De l’escala local a la global

La riquesa d’espècies no és homogènia en l’espai i normalment presenta diferències quan comparem diferents llocs. Aquestes diferències solen respondre a gradients d’un o més factors que creen patrons espacials de biodiversitat i depenen de l’escala. A escala local, els patrons de diversitat depenen de l’extensió de l’hàbitat (la relació espècies-àrea), de la productivitat, de l’hostilitat de l’entorn, de la freqüència i intensitat de les pertorbacions o de la reserva regional d’espècies. La diversitat regional es pot veure afectada per l’heterogeneïtat ambiental (la qual augmenta la dissimilitud), encara que també pot actuar a escales espacials majors o menors, i la connectivitat entre diferents hàbitats. Finalment, a escala global, els patrons de diversitat es corresponen amb la latitud, l’altitud o la profunditat, encara que aquests factors representen una o més variables ambientals (productivitat, àrea, aïllament o hostilitat).
Paraules clau: riquesa d’espècies, relació espècies-àrea, productivitat, gradient latitudinal de biodiversitat.
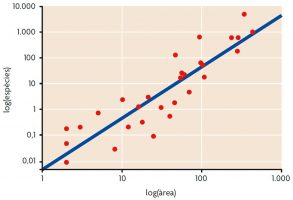
Figura 1. Relació espècies-àrea de les plantes vasculars de l’arxipèlag de les Galápagos: un increment de l’àrea de mostra resulta en un increment del nombre d’espècies trobades. Elaborada a partir de Connor i McCoy, 2000 / Encyclopedia of Biodiversity
Per què alguns llocs són més diversos que altres?
Una ràpida ullada al nostre entorn pròxim hauria de ser prou perquè ens adonàrem que la distribució d’espècies no és homogènia en l’espai i que alguns llocs contenen més espècies que altres amb la mateixa superfície. De fet, les espècies presents en un determinat lloc són el resultat de diversos processos que operen a diferents escales, com les barreres geogràfiques, les limitacions ambientals o les interaccions biòtiques que determinen la composició de les comunitats ecològiques. Tots aquests processos defineixen la composició d’espècies en cada comunitat: el nombre i la identitat d’espècies (o la biodiversitat) en una zona determinada. Quins factors específics influeixen en aquesta distribució heterogènia de la diversitat en l’espai? Aquesta pregunta ha despertat l’interès dels naturalistes des que van començar a explorar el món. Amb el pas del temps van observar que la biodiversitat de molts grups taxonòmics seguia patrons espacials predicibles, per la qual cosa la distribució de la biodiversitat a l’espai es podia explicar gràcies a certs factors, i els patrons observats es repetien en moltes regions del món.
«Els naturalistes van observar que la biodiversitat de molts grups taxonòmics seguia patrons espacials predicibles»
Un dels primers naturalistes que va descriure la relació entre els organismes i els seus entorns va ser Alexander von Humboldt (1769-1859), qui es va centrar principalment en l’efecte de factors geogràfics com el clima sobre diferents tàxons. El treball de Humboldt en la seua expedició científica a Amèrica va inspirar profundament altres naturalistes com Charles Darwin (1809-1882) o Alfred Russel Wallace (1823-1913) a dur a terme les seues exploracions del món. Totes aquestes expedicions van conduir a la posterior cerca de patrons de distribució d’espècies i de diversitat a gran escala. Des de llavors, s’han proposat moltes hipòtesis per a contestar aquesta pregunta, però els factors específics que subjauen als gradients de diversitat continuen essent controvertits. A més, encara que alguns dels patrons poden actuar a diferents escales, la majoria en depenen; per tant, en funció de l’escala espacial que s’estudie, els factors relacionats amb els gradients de diversitat canviaran.
«Probablement els patrons de biodiversitat a escala local van ser els primers a ser descrits, com A resultat d’observacions de camp bàsiques i senzilles»
És important subratllar que molts estudis que analitzen patrons espacials de biodiversitat utilitzen el nombre d’espècies (la riquesa d’espècies) com a variable de resposta. Recentment, els biòlegs han començat a centrar-se en altres aspectes de la biodiversitat com la diversitat genètica o funcional. No obstant això, per una qüestió de simplicitat, centrarem aquest repàs només en la diversitat d’espècies. Per tant, d’ara en avant utilitzarem de manera indistinta els termes biodiversitat i riquesa d’espècies. Tot seguit discutirem amb més detall els patrons de biodiversitat a diferents escales espacials (resumits en la Taula 1).
Patrons locals
Probablement els patrons de biodiversitat a escala local van ser els primers a ser descrits, com a resultat d’observacions de camp bàsiques i senzilles. Un dels primers patrons a ser observats va ser la relació entre nombre d’espècies i àrea: com major és l’àrea mostrejada, major és el nombre d’espècies que alberga (Figura 1). Aquest patró s’ha observat en tot el món, tant en entorns terrestres com en marins. La primera descripció de la relació espècies-àrea es va realitzar en el segle XIX, quan H. C. Watson va observar en un comtat d’Anglaterra que el nombre d’espècies de plantes en una mostra es doblava en incrementar deu vegades l’àrea estudiada (Connor i McCoy, 2000). Després, Arrhenius va quantificar aquesta relació per primera vegada en 1921 mitjançant la funció S = cAz (on S és el nombre d’espècies, A és l’àrea i c i z són constants). Les constants c i z serveixen per a establir comparacions entre diferents àrees d’estudi.

Figura 2. Un dels patrons de la relació entre espècies i àrea indica que les illes més grans d’un arxipèlag alberguen un nombre més gran d’espècies. Aquesta és una de les bases de la teoria de biogeografia d’illes de MacArthur i Wilson. En la imatge, foto satèl·lit de l’arxipèlag de Hawaii (EUA). /Jacques Descloitres, MODIS Land Rapid Respon-se Team, NASA GSFC Figura 3. Hi ha factors que determinen els patrons de biodiversitat i limiten el nombre d’espècies; per exemple, l’hostilitat de l’ambient, com ocorre en hàbitats amb temperatures extremes. En la imatge, la vall desèrtica coneguda com Wadi Rum, a Jordània. / Martino Pietropoli
Rosenzweig (1997) considera que en realitat les corbes espècies-àrea es podrien dividir en quatre patrons distints. Els dos primers depenen de l’extensió de l’àrea mostrejada: menuda o gran. Un tercer considera la macroescala (províncies biogeogràfiques), en la qual l’increment d’espècies no es relaciona amb la immigració d’espècies des d’altres àrees, sinó amb els processos d’especiació, que actuen a una escala temporal més lenta. I el quart patró és la base de la teoria de biogeografia d’illes de MacArthur i Wilson, que indica que les illes més grans d’un arxipèlag alberguen un nombre més gran d’espècies (Figura 2). S’utilitzen dos arguments principals per a explicar la relació entre nombre d’espècies i àrea: primer, que les àrees més grans poden mantenir poblacions més grans i, per tant, les espècies tenen menys probabilitats d’extingir-se; i segon, que els hàbitats amb superfícies més grans són més heterogenis (explicat més avant a escala regional).
| Escala | Factor |
| Local | Àrea |
| Productivitat de l’ecosistema | |
| Hostilitat de l’entorn | |
| Nivell de pertorbació | |
| Reserva regional d’espècies | |
| Regional | Heterogeneïtat ambiental |
| Connectivitat | |
| Global | Latitud |
| Altitud (en muntanyes) | |
| Profunditat (en ecosistemes marins) |
Taula 1. Resum dels diferents factors que creen patrons espacials de diversitat i les escales en què operen: local, regional i global. En alguns casos, el factor no és la causa que subjau al gradient de diversitat, sinó que és una suma de diferents variables la que crea el patró (per exemple la latitud; vegeu el text).
Un altre patró que s’ha estudiat bastant és la relació entre riquesa d’espècies i productivitat en un ecosistema. La productivitat és el ritme a què es produeix biomassa en una àrea determinada. És una mesura de l’aportació energètica a un ecosistema (normalment estimada mitjançant la precipitació, evapotranspiració o aportació de nutrients). Originàriament es pensava que els recursos tendien a ser il·limitats en els nivells més alts de productivitat, la qual cosa permetia la presència d’un nombre més gran d’espècies. Aquest patró s’observa principalment a escala global, on les regions biogeogràfiques amb més aportació energètica generalment contenen més espècies (vegeu l’apartat de patrons globals). A escales locals s’han observat diferents tipus de relacions: positiva, negativa, en forma de U i en forma de campana (Mittelbach et al., 2001). Aquesta última la trobem quan la biodiversitat és major amb nivells intermedis de productivitat, i és bastant freqüent en la naturalesa com per haver motivat nombrosos estudis que intenten explicar les seues causes. En la primera part del gradient de productivitat (productivitat baixa o mitjana) la diversitat d’espècies augmenta a mesura que augmenta la disponibilitat de recursos. No obstant això, la disminució en la riquesa d’espècies que es produeix a partir d’un cert nivell de productivitat (la paradoxa de l’enriquiment) no està tan clara. S’han suggerit diverses hipòtesis, com l’augment de l’exclusió competitiva quan hi ha nivells alts de productivitat o el canvi en el recurs limitant per a les plantes, que passa de ser els nutrients a ser la llum (Tilman i Pacala, 1993). No obstant això, cap hipòtesi explica la variació en forma i magnitud de la relació entre productivitat i diversitat, i factors com l’escala espacial, els tàxons estudiats, el tipus d’hàbitat (terrestre o aquàtic) o la intensitat de la depredació (coexistència mediada per l’explotador) poden ser rellevants per a explicar aquesta disparitat.
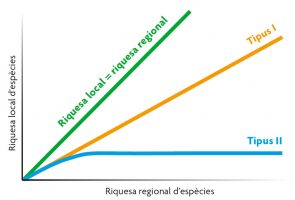
Figura 4. Relació entre riquesa d’espècies local i regional. La figura mostra la línia teòrica en què la riquesa local és igual que la riquesa regional, així com els dos tipus diferents de respostes: de tipus I, una relació positiva, quan la composició de parcel·les locals depèn de la riquesa regional, i de tipus II, quan les parcel·les locals tenen característiques específiques que impedeixen l’entrada d’espècies des de la reserva regional d’espècies. Elaborada a partir de Gaston, 2000 / Nature
Encara que tant l’àrea com la productivitat afavoreixen la coexistència de nombroses espècies, altres factors determinen els patrons de biodiversitat limitant-les en nombre. Per exemple, l’hostilitat de l’entorn (hàbitats molt alcalins o àcids, o amb temperatures extremes) selecciona les espècies que hi poden sobreviure, organismes amb adaptacions molt específiques (Figura 3). No obstant això, per a complicar més les coses, potser aquests hàbitats també compartesquen característiques comunes que podrien reduir la riquesa d’espècies, com una àrea petita o un cert grau d’aïllament (per exemple, aigües termals o cims de muntanyes). Això dificulta la possibilitat de separar l’efecte de tots aquests factors en la diversitat local. Una altra hipòtesi proposada per a explicar els gradients de diversitat és la hipòtesi de la pertorbació intermèdia (Connell, 1978), que suggereix que els hàbitats amb nivells alts i baixos de pertorbació contenen poques espècies. Per tant, la màxima diversitat es trobaria en els nivells intermedis de pertorbació (Connell, 1978), perquè aquests impedeixen la dominància de bons colonitzadors (abundants amb nivells alts de pertorbació) i competidors (amb nivells baixos). Encara que la primera demostració empírica es va realitzar en roques intermareals sotmeses a diferents graus d’intensitat de tempestes (Sousa, 1979), els estudis realitzats amb altres sistemes no han aconseguit trobar proves que abonen la hipòtesi de la pertorbació intermèdia.
«L’heterogeneïtat ambiental és un dels principals factors que creen un gradient de diversitat a escala regional»
Finalment, l’extensió de la reserva regional d’espècies és rellevant per a la diversitat local. Què ocorre en comunitats locals en què hi ha un gradient en la riquesa regional d’espècies? En teoria es podrien observar dues respostes distintes (Figura 4). La primera la trobem quan la composició de les comunitats locals depèn en gran manera de la reserva regional d’espècies (espècies que arriben i colonitzen les comunitats locals). En aquest cas, trobarem una relació lineal positiva: a mesura que augmente la diversitat regional, també ho farà la diversitat local (corba de tipus I). La segona es produirà quan les comunitats locals presenten algunes característiques que limiten l’entrada indiscriminada d’espècies de la regió (per exemple, competència o depredació). En aquest cas, la riquesa local se saturarà amb la riquesa regional perquè el nombre d’espais ecològics en els hàbitats locals és limitat (corba de tipus II). Els estudis empírics han conclòs que la resposta de tipus I és la més comuna en la naturalesa.

Patrons regionals
Es considera que una regió inclou un gran nombre d’hàbitats i comunitats, i sovint s’hi fa referència com l’àrea des de la qual les espècies colonitzen comunitats locals. Per tant, un dels principals factors que creen un gradient de diversitat a escala regional és l’heterogeneïtat ambiental: si l’entorn de la regió és homogeni, les seues comunitats probablement continguen les mateixes espècies, la qual cosa resulta en una riquesa regional d’espècies baixa.1 Al contrari, una variació àmplia de les condicions ambientals en diferents hàbitats d’una regió permetrà la presència de comunitats diverses (molt dissimilars)2 i una riquesa regional d’espècies alta (Figura 5). La relació positiva entre heterogeneïtat ambiental i riquesa d’espècies s’ha demostrat de manera empírica en diferents tàxons, ecosistemes i escales espacials (Stein, Gerstner i Kreft, 2014). A més de la varietat d’hàbitats en una regió, aquest patró també opera a escales espacials menors (heterogeneïtat de microhàbitats a escala local) i majors (gradients d’hàbitats a escala global).
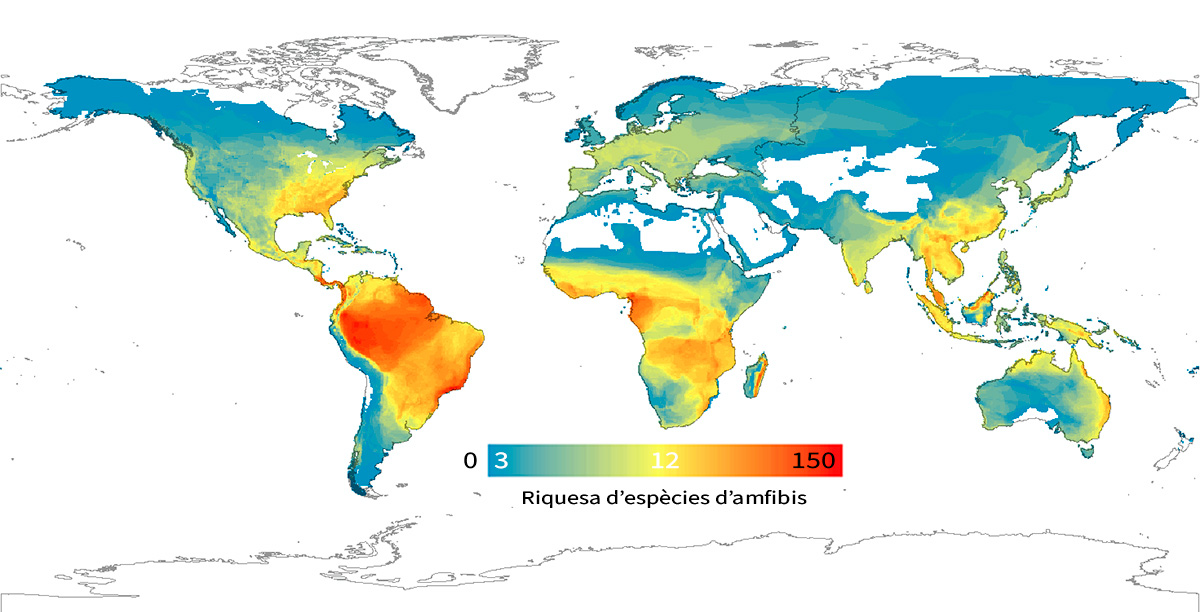
Figura 6. Gradient latitudinal de diversitat en amfibis, que inclou 6.117 espècies en cel·les de 5 graus de latitud. El nombre d’espècies és major en els tròpics i disminueix gradualment cap als pols. El gradient de colors indica una variació que va de 3 espècies per cel·la (blau), a 12 (groc) i a 150 (roig) aproximadament. Font: Pyron i Wiens, 2013. Publicat per la Royal Society. Tots el drets reservats.
En les últimes dècades, amb el creixent interès en les metacomunitats (conjunt de comunitats locals connectades per dispersió), el flux d’individus entre parcel·les locals s’ha començat a tenir en compte. Un dels factors importants que afecten aquest flux és el grau de connectivitat de les parcel·les locals, determinat per la localització de cada parcel·la en el paisatge (Borthagaray, Pinelli, Berazategui, Rodríguez-Tricot i Arim, 2015). Així, depenent de la configuració de la xarxa de la metacomunitat, així com de la capacitat de dispersió de les espècies i la grandària de cadascuna, la riquesa d’espècies local i regional pot variar. Per exemple, en el cas d’organismes amb taxes de dispersió baixes, quan les parcel·les estan aïllades, la diversitat local seria baixa, però la regional seria alta. Al contrari, quan els organismes es dispersen fàcilment i les parcel·les estan molt interconnectades, la diversitat local seria alta i la regional baixa.

Figura 7. Il·lustració de l’«efecte de domini mitjà» amb llapis per a mostrar una de les possibles explicacions del gradient latitudinal de diversitat (la disminució de diversitat des de l’equador als pols o en qualsevol àrea amb uns límits clars): per atzar, el centre d’una caixa plena de llapis de diferents grandàries contindrà més llapis (o més parts) que els extrems. Elaborada a partir de Mittelbach, 2012 / Community Ecology
Patrons globals
Com ja s’ha indicat, les primeres evidències dels gradients de biodiversitat a gran escala ens les van proporcionar les primeres expedicions europees al Nou Món després del segle XVIII. En aquestes exploracions, els naturalistes van ser enviats a descriure les espècies exòtiques del lloc per a aconseguir nova informació sobre la misteriosa i exòtica naturalesa d’aquest nou continent. Molts d’ells van quedar fascinats per la gran diversitat d’espècies (i les seues formes, colors i comportaments) dels tròpics en comparació amb les ben conegudes regions europees. Aquestes observacions van conduir a la descripció del patró de biodiversitat a gran escala més famós: el gradient latitudinal de diversitat, caracteritzat per una disminució en la riquesa d’espècies des de l’equador cap a latituds majors (al nord o al sud) (Figura 6). Aquesta disminució en la diversitat és lleugerament asimètrica entre els dos hemisferis, amb una corba més pronunciada en l’hemisferi nord. El gradient latitudinal de diversitat s’ha documentat en una varietat de tàxons animals i vegetals de diferents ecosistemes (marins i terrestres; Hillebrand, 2004). No obstant això, hi ha algunes excepcions. Per exemple, els macròfits aquàtics representen un dels pocs grups taxonòmics que mostren un gradient latitudinal de diversitat invers, amb comunitats més riques en latituds més altes.
A més, aquest gradient no s’observa només en espècies actuals, sinó també en el registre fòssil (s’ha observat sobretot en alguns tàxons d’invertebrats marins, com els braquiòpodes o els foraminífers). No obstant això, la latitud en si mateixa no és la causa subjacent d’aquest gradient de biodiversitat. Aquesta correlació podria explicar-se amb molts factors ambientals. Fins ara s’han suggerit molts mecanismes diferents i, probablement, la combinació d’alguns d’aquests afecte la variació en la diversitat al llarg del gradient latitudinal. Mittelbach (2012) resumeix totes les possibles hipòtesis en quatre grups: ecològiques, històriques, evolutives i el model nul. Les hipòtesis ecològiques es basen principalment en la relació entre productivitat (l’aportació energètica i la disponibilitat de recursos) i una major abundància d’individus, la qual cosa redueix el risc d’extinció. Les hipòtesis històriques defensen que els tròpics són més antics i que tenen una major extensió geogràfica (en el passat i en l’actualitat) i unes condicions climàtiques més estables en el temps, per la qual cosa han tingut més temps per a la diversificació. Les hipòtesis evolutives se centren en les taxes més altes de diversificació en els tròpics. I, finalment, el model nul es basa en l’«efecte de domini mitjà»: la idea que si els rangs de distribució d’espècies es localitzen a l’atzar al llarg del gradient latitudinal és més probable que un nombre més gran de distribucions d’espècies se solapen en el mig (en l’equador). Podem veure-ho fàcilment amb una caixa plena de llapis de diferents grandàries en què, per atzar, la majoria de llapis estaran en el centre de la caixa (Figura 7).
«El gradient latitudinal de diversitat no s’observa només en espècies actuals, sinó també en el registre fòssil»
Encara que el gradient latitudinal de diversitat és el patró més apassionant de l’ecologia, s’han descrit altres patrons globals, com els gradients d’elevació en sistemes muntanyosos i els gradients de profunditat en entorns marins. Com la latitud, aquestes dues variables no són la causa que subjau als patrons de riquesa d’espècies. En aquests casos, les condicions ambientals, més hostils a mesura que s’avança en el gradient d’altitud i profunditat, així com la baixa productivitat, l’aïllament o les àrees reduïdes (per exemple, els pics més alts), poden contribuir a la disminució en la riquesa d’espècies.
Què podem aprendre dels patrons espacials de biodiversitat?
Hem presentat els patrons de biodiversitat més estudiats a diferents escales espacials. Alguns d’ells presenten una relació clara i generalitzada amb la diversitat, mentre que en altres casos la resposta és qüestionable i necessita ser investigada amb més profunditat. No obstant això, hem de subratllar que la majoria d’aquests patrons s’han establit amb importants llacunes de coneixement en allò referent a la taxonomia i distribució d’espècies, especialment a escala global (parlem dels coneguts com a «dèficit de Linné» i «dèficit de Wallace», respectivament; Hortal et al., 2015). Aquestes llacunes de coneixement reflecteixen no sols les diferències en els esforços realitzats per a investigar en diferents parts del món, sinó també la desigualtat en els tàxons estudiats. Per exemple, els conjunts de dades sobre espècies de vertebrats (aus o mamífers) i alguns grups de plantes (arbres) són molt més complets que els d’invertebrats o altres tàxons més petits. De la mateixa manera, els hàbitats aquàtics s’han estudiat molt poc, a pesar de la seua alta i desproporcionada contribució a la diversitat global, i encara hi ha una certa incertesa sobre si la biodiversitat en els entorns aquàtics segueix els mateixos patrons que en els hàbitats terrestres (Siqueira, Bini, Thomaz i Fontaneto, 2015). Eliminar aquestes llacunes ajudarà a aclarir la ubiqüitat dels patrons espacials i les seues excepcions (si les haguera).
«Les primeres proves dels gradients de biodiversitat a gran escala ens les van proporcionar les expedicions europees al nou món després del segle XVIII»
Els gradients espacials de biodiversitat i les seues causes no són rellevants només des d’un punt de vista naturalista; també tenen implicacions importants per a la conservació de la biodiversitat. Per exemple, els punts calents de diversitat de tot el món es poden establir seguint aquests gradients. A escala global, la importància dels tròpics per a la biodiversitat és indiscutible, per la qual cosa hauríem de fer esforços importants centrats en aquestes àrees. A escala local i regional, les àrees protegides haurien de dissenyar-se considerant els factors ambientals que maximitzen la preservació de la biodiversitat. Així, hauríem de tenir en compte factors com la delimitació de grandària, la inclusió de diferents hàbitats per a augmentar l’heterogeneïtat o considerar la connectivitat o aïllament de parcel·les. Finalment, després del treball experimental i de camp de les últimes dècades del segle XX, els beneficis actuals de la biodiversitat han estat àmpliament reconeguts. A més de béns inestimables (aliment, aigua o medicines), la biodiversitat està correlacionada amb el funcionament i els serveis d’un ecosistema: millora la productivitat, el cicle de nutrients, l’estabilitat de l’ecosistema o la resistència a espècies invasores. Per tant, conèixer i preservar les àrees amb alta diversitat contribuirà a mantenir aquests beneficis.
1. El nombre d’espècies que es troba en una regió rep el nom de diversitat gamma. (Tornar)
2. La similitud o dissimilitud entre comunitats locals rep el nom de diversitat beta. Mesura la diferència en la composició d’espècies entre dos o més hàbitats d’una regió. (Tornar)
Referències
Borthagaray, A. I., Pinelli, V., Berazategui, M., Rodríguez-Tricot, L., & Arim, M. (2015). Effects of metacommunity networks on local community structures: From theoretical predictions to empirical evaluations. En A. Belgrano, G. Woodward, & U. Jacob (Eds.), Aquatic functional biodiversity: An eco-evolutionary approach (pp. 75–111). Londres/San Diego, CA/Waltham, MA/Oxford: Academic Press (Elsevier).
Connell, J. H. (1978). Diversity in tropical rain forests and coral reefs. Science, 199, 1302–1310. doi: 10.1126/science.199.4335.1302
Connor, E. F., & McCoy, E. D. (2000). Species-area relationships. En S. A. Levin (Ed.), Encyclopedia of biodiversity (pp. 397–411). Nova York, NY: Elsevier. doi: 10.1016/B0-12-226865-2/00252-2
Hillebrand, H. (2004). On the generality of the latitudinal diversity gradient. The American Naturalist, 163(2), 192–211. doi: 10.1086/381004
Hortal, J., De Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M., & Ladle, R. J. (2015). Seven shortfalls that beset large-scale knowledge of biodiversity. Annual Review of Ecology, Evolution, and Systematics, 46, 523–549. doi: 10.1146/annurev-ecolsys-112414-054400
Mittelbach, G. G. (2012). Community ecology. Sunderland, MA: Sinauer Associates.
Mittelbach, G. G., Steiner, C. F., Scheiner, S. M., Gross, K. L., Reynolds, H. L., Waide, R. B., … Gough, L. (2001). What is the observed relationship between species richness and productivity? Ecology, 82(9), 2381–2239. doi: 10.2307/2679922
Rosenzweig, M. L. (1997). Species diversity in space and time. Cambridge: Cambridge University Press.
Siqueira, T., Bini, L. M., Thomaz, S. M., & Fontaneto, D. (2015). Biodiversity analyses: Are aquatic ecologists doing any better and differently than terrestrial ecologists? Hydrobiologia, 750, 5–12. doi: 10.1007/s10750-014-2071-6
Sousa, W. P. (1979). Disturbance in marine intertidal boulder fields: The non-equilibrium maintenance of species diversity. Ecology, 60, 1225–1239. doi: 10.2307/1936969
Stein, A., Gerstner, K., & Kreft, H. (2014). Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecology Letters, 17(7), 866–880. doi: 10.1111/ele.12277
Tilman, D., & Pacala, S. (1993). The maintenance of species richness in plant communities. En R. E. Ricklefs & D. Schluter (Eds.), Species diversity in ecological communities (pp. 13–25). Chicago: The University of Chicago Press.
Aquest treball ha rebut el suport de CAPES (Coordenação d’Aperfeiçoamento de Pessoal de Nivell Superior), del Ministeri d’Educació de Brasil.




