

Història i realitat del gènere ‘Homo’
Què és i per què el considerem com a tal?
Els paleantropòlegs que s’ocupen d’investigar com s’organitza la naturalesa en espècies i com hauríem de denominar-les solen ser acusats de «limitar-se a discutir sobre noms». Aquesta idea implica que la taxonomia bàsica és una tasca administrativa avorrida que s’hauria de solucionar ràpidament, o fins i tot que s’hauria d’ignorar, per a poder arribar ràpidament a les qüestions realment interessants sobre l’evolució humana. No obstant això, la realitat és que mai entendrem la trama d’aquesta intricada obra que és l’evolució humana si no identifiquem amb precisió els actors que hi van participar. Aquest article revisa la formació del concepte actual del gènere Homo, que és summament vague, com a rerefons per a instar a realitzar una definició més racional.
Paraules clau: taxonomia, gèneres, gènere Homo, paleoantropologia.
El gènere ‘Homo’: Enfocament de la qüestió
La primera qüestió que sorgeix en definir el gènere Homo és la naturalesa de la mateixa unitat. Avui dia, s’accepta majoritàriament que en principi els gèneres no haurien de ser només monofilètics, sinó holofilètics. És a dir, un gènere hauria de contenir totes les espècies descendents d’un avantpassat comú concret, i cap altra. Però no hi ha una regla, ni tan sols una d’empírica, que diga quantes espècies hauria d’haver-hi en cada gènere, o com de remot pot arribar a ser aquest avantpassat comú. Alguns gèneres actuals ben establerts són rics en espècies, mentre que altres en tenen moltes menys i alguns inclouen una única espècie. En part, aquesta varietat es dóna perquè alguns gèneres semblen més intrínsecament disposats que altres a la diversificació; però en part també és degut a una flexibilitat taxonòmica que reconeix que no hi ha fronteres «naturals» per a un gènere (com sí que n’hi ha, almenys en teoria, per a les espècies).
«Un gènere hauria de contenir totes les espècies descendents d’un avantpassat comú concret, i cap altra»
Afortunadament, en el món dels éssers vius, els gèneres tendeixen a ser fàcilment recognoscibles com a unitats Gestalt.1 Això és degut al fet que les distàncies morfològiques entre gèneres relacionats solen ser molt més grans que els que separen espècies del mateix gènere, ja que a mesura que s’expandeix la xarxa, les diferències s’acumulen de manera natural, encara que a un ritme irregular. Els judicis taxonòmics són inherentment més complicats utilitzant registres fòssils que éssers vius, no sols perquè la informació disponible és necessàriament més limitada, sinó també perquè les proves que sí que tenim tendeixen a ser més aviat incompletes i estan disperses en el temps i l’espai.
La nostra comprensió actual de cada branca de la ciència està profundament condicionada per la història particular de cada camp. I en el cas de la paleoantropologia és especialment cert que la creença actual és producte –fins a un punt alarmant– de la seqüència completament arbitrària en què es van descobrir els components del registre fòssil conegut.
Les nostres idees paleoantropològiques actuals també s’han vist profundament afectades en l’últim parell de segles per les enormes fluctuacions en les nostres idees sobre el que són i haurien de ser els gèneres. A causa d’aquesta realitat, aquestes consideracions sobre el gènere Homo són principalment històriques. Perquè quasi amb tota seguretat, si demà es descobrira el registre fòssil homínid al complet en compte d’anar trobant elements a poc a poc al llarg de segle i mig, ho veuríem de manera molt diferent de la manera com ho veiem avui.
Primeres nocions del gènere ‘Homo’
El concepte original del gènere Homo, encunyat per Linné (Linnaeus, 1758), es podia comparar estretament amb la superfamília Hominoidea actual, ja que comprenia no sols Homo sapiens sinó també Homo troglodytes, una espècie que incloïa tant ximpanzés com orangutans. La noció que Homo sapiens podria haver compartit gènere amb parents extints avui dia no va aparèixer fins a 1864, quan William King va classificar com a Homo neanderthalensis un esquelet fòssil parcial clarament antic que s’havia trobat a la vall (Thal en alemany) de Neander a Alemanya (King, 1864). Durant tres dècades, però, no es va descriure cap nova forma extinta d’Homo, principalment perquè la discussió sobre els orígens biològics de la humanitat va continuar en absència de qualsevol fòssil, a part d’uns pocs que s’assemblaven als neandertals. Encara que no tots ho veien com un obstacle.
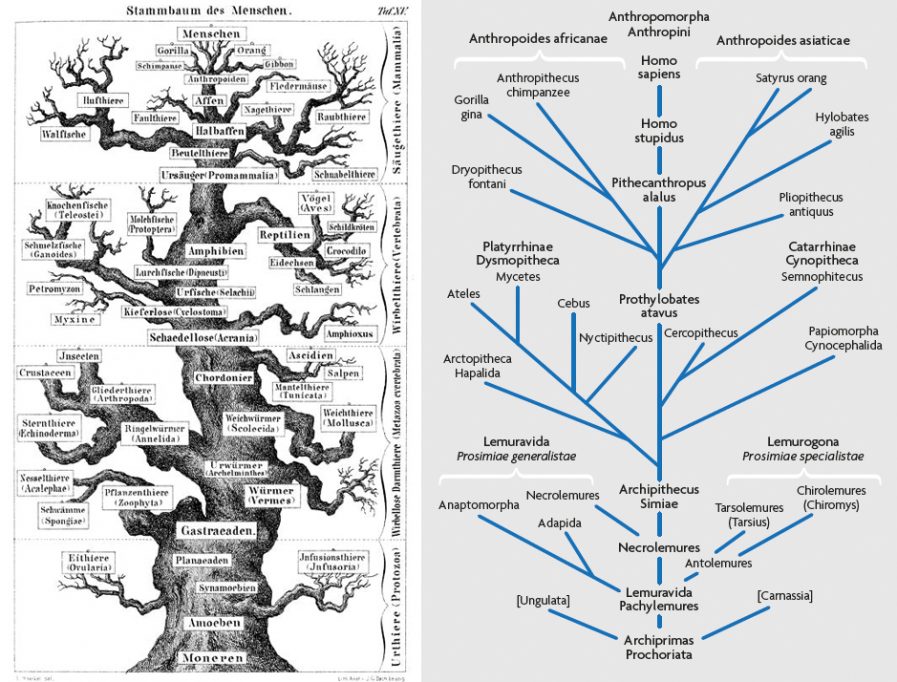
La Figura 1 mostra la noció d’Ernst Haeckel de l’arbre de la vida, publicada en 1868, quan, a part dels neandertals, amb prou feines hi havia un registre fòssil humà. El dibuix de Haeckel representa la clàssica imatge d’un arbre amb un tronc estret i branquillós i Homo sapiens (Menschen) al cimal. En una filogènia dels primats posterior i més detallada (Figura 2), Haeckel va anar fins i tot més lluny, encara que en aquell moment ell només podia imaginar el que la història fòssil de la humanitat revelaria. Però sabia que devia haver-hi connexions anteriors entre els éssers humans i la resta de la naturalesa, per la qual cosa va inserir en la seua filogènia dues espècies hipotètiques, Homo stupidus (que no el neandertal, si és el que estàveu pensant) i Pithecanthropus alalus, connectats per ascendència i descendència directa i lineal (Haeckel, 1868).
Per això en 1891, quan Eugene Dubois va descobrir a Trinil (Java) la calota cranial2 de l’espècie fòssil d’homínids que més tard es coneixeria com Pithecanthropus erectus, li va semblar perfectament lògic considerar l’espècimen com un avantpassat lineal d’Homo sapiens, ja que tenia un cervell petit però clarament humà en un cert sentit. Aquesta increïble troballa semblava encaixar perfectament en el constructe de Haeckel; fins a tal punt que Dubois va utilitzar per al seu fòssil el nom que Haeckel donava al gènere, Pithecanthropus (“simi-home”). Aquesta posició intermèdia es reflecteix en el diagrama filogenètic de Dubois de 1896 (Figura 3, vegeu Theunissen, 1988). Gràcies als esforços de Dubois, però també de molts altres acadèmics influents, com l’anatomista alemany Gustav Schwalbe, a les acaballes del segle XX hi havia un ampli consens i tothom es pensava que Pithecanthropus erectus de Dubois era l’espècie ancestral que havia donat lloc a Homo sapiens.
«La noció que ‘Homo sapiens’ podria haver compartit gènere amb parents extints avui dia no va aparèixer fins a 1864»
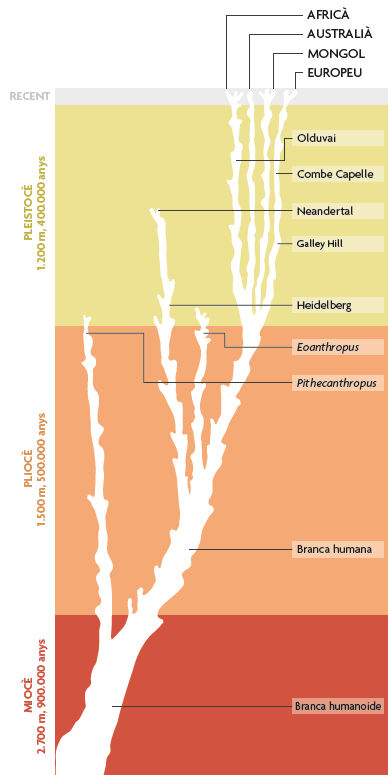
Però sorprenentment prompte, i potser de manera inevitable, les interpretacions van començar a canviar. Al començament de la segona dècada del segle XX, el Pithecanthropus de Dubois s’havia tornat, una vegada més, clarament marginal, com es veu en la Figura 4, que mostra un diagrama filogenètic humà publicat per l’anatomista britànic Sir Arthur Keith (1915). En aquesta concepció, quasi tots els fòssils humans o grups de fòssils coneguts en aquell moment es veien com a parts, o com el final, de llinatges terminals. És significatiu que la majoria dels qui escrivien sobre el registre fòssil humà durant aquell període eren anatomistes humans com Keith: científics molt versats en els petits detalls de variació d’Homo sapiens, però amb poc interès en els debats sobre mecanismes evolutius entre genetistes, embriòlegs, naturalistes i altres experts oberts durant les dècades posteriors al redescobriment en 1900 dels principis de la genètica mendeliana. Els paleoantropòlegs d’aquest període semblen haver acceptat en general la noció darwinista d’evolució com a «descendència amb modificació», i deixen que altres es preocupen dels processos subjacents. Es limitaven a ocupar-se d’examinar els resultats. No obstant això, a diferència de l’obligació a què s’enfrontaven els anatomistes comparatius, això no va implicar necessàriament haver de classificar la diversitat dels éssers vius. Com a resultat, i en el que concernia als paleoantropòlegs, els noms, fins i tot les denominacions zoològiques formals, eren poc més que identificadors convenients per a fòssils individuals, més que no una forma de perfilar unitats en la naturalesa. Sorprenentment, això s’aplica tant als gèneres com a les espècies: per a la majoria de paleoantropòlegs d’aquest període, el nostre era un gènere que contenia únicament Homo sapiens i homínids extints amb una grandària cerebral semblant, la qual cosa sovint incloïa els neandertals en el gènere Homo, però no gaires espècies més.
Per a quan va arribar la Segona Guerra Mundial, qualsevol patró discernible en un registre fòssil que s’expandia de manera constant havia quedat enfosquit per un garbuix de noms informals i genèrics. I sense importar la nomenclatura preferida, l’arbre familiar homínid típic durant el període d’entreguerres va continuar, a l’estil de Keith, representant l’evolució homínida com un conjunt abundant de branques terminals.
La síntesi evolutiva
El moviment conegut com a síntesi evolutiva va nàixer durant les dècades de 1930 i 1940 a partir d’una fusió teòricament incòmoda (encara que atractiva en termes reduccionistes) de la genètica de poblacions de base matemàtica i la teoria de l’especiació. En última instància, va reduir la complexitat del procés evolutiu a una simple qüestió de canvis graduals en les freqüències amb què apareixien els gens en els llinatges d’organismes en evolució, guiats per la influència de la selecció natural. Especialment en les seues versions més «dures», la síntesi mostrava que cada espècie es transformava gradualment en la seua successora al llarg d’extensos períodes de temps. L’aparició dels gèneres i tàxons superiors es veia simplement com a conseqüència passiva d’aquest tipus de transicions repetides.
«Per a la majoria, el nostre gènere contenia únicament ‘Homo sapiens’ i homínids extints amb una grandària cerebral semblant»
Un dels proselitistes més influents de la síntesi evolutiva va ser l’ornitòleg Ernst Mayr, qui va ser més responsable que qualsevol altre d’acostar finalment la teoria evolutiva a la paleoantropologia. En 1950, Mayr va publicar un article que reescrivia literalment les normes amb què havien d’operar els paleoantropòlegs. Per a entendre la importància de la revolució provocada per Mayr, cal recordar que els relativament escassos fòssils d’homínids coneguts en aquell moment es caracteritzaven per una profusió de noms de gèneres i espècies. Expeditivament, Mayr va reduir uns quinze noms de gèneres a un de sol. És més, va reduir literalment dotzenes de noms d’espècies a únicament tres, i va crear la següent classificació de la família homínida:
Família Hominidae
Gènere Homo
Espècie Homo transvaalensis [australopitecs]
Espècie Homo erectus
Espècie Homo sapiens [incloent-hi els neandertals]
Finalment, el gènere Homo tornava a estar al bell mig de la paleoantropologia. De fet, la visió de Mayr comprenia la família Hominidae al complet. És més, Mayr va ubicar les seues tres espècies del gènere Homo en un únic llinatge evolutiu, tornant a la linealitat pura de l’enfocament haeckelià. En proposar aquesta excessiva simplificació, Mayr va tenir molt poca consideració pel suposat enfocament «no biològic» dels paleoantropòlegs anteriors a la guerra, als quals directament va reprovar per no pensar en termes de variabilitat dins de les poblacions biològiques. Sorprenentment, almenys en l’entorn de parla anglesa, els paleoantropòlegs van acceptar la contundent crítica de Mayr amb una humilitat sorprenent. De fet, es van precipitar a unir-se al carro de l’evolucionisme unilineal, possiblement perquè sempre havien sigut conscients subliminarment del buit teòric que ocupava el centre de la seua disciplina. Tan profunds van ser els efectes de l’atac de Mayr que, en menys d’una dècada, ningú gosava utilitzar noms de gèneres o espècies en absolut.
La preocupació de Mayr amb la variació intraespecífica era, per descomptat, vàlida. I s’ajustava còmodament a la tradició anatòmica humana de documentar la variació d’Homo sapiens. Però el nou enfocament també significava que ningú es preocupava en absolut de les sinapomorfies morfològiques3 que podrien haver mantingut el renascut gènere Homo com una unitat. En canvi, es van crear diagrames com el de la Figura 5, a partir d’un article clàssic escrit pel paleoantropòleg nord-americà Clark Howell (1957).
«La síntesi evolutiva va nàixer a partir de la fusió de la genètica de poblacions de base matemàtica i la teoria de l’especiació»
En el diagrama de Howell els participants en el joc evolutiu són tots fòssils individuals, majoritàriament continguts en una única massa filogenètica sense forma que no evidencia de cap manera la naturalesa de la diversitat entre els homínids, o el que podria unir el gènere Homo. Anys després, els paleoantropòlegs continuaven publicant de manera rutinària filogènies minimalistes d’aquest tipus, amb una soca principal àmplia, la dels homínids, que donava lloc, com a màxim, a un sol llinatge secundari, aberrant i efímer: els robustos australopitecs, els quals fins i tot Mayr (1953) es va veure obligat a deixar de costat després d’un exabrupte de John Robinson (1951). En aquests constructes tan lineals, les espècies –i per extensió, els gèneres– simplement es classificaven de manera inconsistent l’una darrere de l’altra al llarg del temps. I la conseqüència d’això era que resultava impossible, quasi per definició, reconèixer espècies i gèneres basant-se en caràcters morfològics derivats (fins i tot encara que algú ho haguera intentat). Els paleoantropòlegs havien oblidat –si alguna vegada ho van comprendre de veritat– que els gèneres, com a agrupacions d’espècies, són tàxons superiors i que, en conseqüència, necessitem definir el gènere Homo, com qualsevol altre clade, basant-nos en les sinapomorfies.
Després de la síntesi
En aquest context, no és sorprenent que la dècada de 1960 també fóra el període en què es va estendre la noció ultraminimalista de l’evolució humana com «una única espècie». Els antecedents d’aquesta noció són estranys i variats. Un d’aquests era Franz Weidenreich, el brillant anatomista i estudiós de l’home de Pequín (Weidenreich, 1939). Com la majoria dels seus contemporanis, Weidenreich es va veure poc afectat per qualsevol teoria sobre com funcionava realment l’evolució, encara que sembla que es va decantar per l’ortogènesi, la noció que l’evolució està, d’alguna manera, guiada per un objectiu. Aquesta idea era un anatema per als altres progenitors de la hipòtesi d’una sola espècie, Theodosius Dobzhansky (1944) i Ernst Mayr (1950); no obstant això, els tres membres del trio se sentien còmodes amb la idea que l’adquisició de la cultura havia ampliat tant el nínxol ecològic de l’ésser humà que, fins i tot per definició, no podria haver existit més d’una espècie humana en un moment donat del temps. I si aquest fóra realment el cas, significaria que, sense importar com diferesquen entre ells en termes anatòmics, tots els fòssils homínids del mateix període haurien de pertànyer necessàriament a la mateixa espècie.
«L’impressionant registre fòssil de què disposem avui dia apunta realment a una diversitat d’espècies en la història dels homínids»
Encara que a mitjan segle ja era difícil basar-se en morfologies fòssils, tal va ser l’impacte de Mayr per a la paleoantropologia que no va ser fins a mitjan dècada de 1970, quan es va demostrar que dos tipus diferents d’homínid havien conviscut en el mateix paisatge del nord de Kenya fa menys de dos milions d’anys, quan els paleoantropòlegs es van veure forçats una vegada més a pensar en termes de diversitat sistemàtica entre els primers homínids. Des d’aquell moment, el registre fòssil humà es va acumular ràpidament i va mostrar que la història de la família humana no responia només al perfeccionament gradual d’adaptacions en un únic llinatge humà principal. Al contrari, la història de la família homínida ha implicat una experimentació evolutiva constant, amb l’aparició regular de noves espècies que competeixen en l’escenari ecològic i moltes vegades s’extingeixen. I lluny de ser el cimal d’un arbre alt i esvelt, com consideraven Haeckel i Mayr, Homo sapiens és només el final d’un branquilló en un arbust exuberant d’una complexitat que es fa més i més evident amb el temps.
En 1993 vaig publicar una filogènia molt conjectural de la família homínida (Tattersall, 1993), que mostra onze espècies d’homínids, i va ser considerada bastant extravagant en aquell moment (Figura 6). Per a quan es va redibuixar el diagrama en 1998, el compte havia ascendit a setze. Curiosament, moltes de les espècies addicionals havien aparegut en la base de l’arbre, cosa que demostrava que el patró de diversitat i d’experimentació evolutiva observat en l’original havia estat present entre els homínids des del començament. En 2002 la imatge s’havia tornat encara més complexa, amb vint espècies que calia encaixar d’alguna forma; i la versió més recent (Figura 7) conté 27 espècies. Els detalls d’aquestes filogènies tan incertes, de la mateixa manera que el nombre exacte d’espècies que es consideren, continuen sent tan discutibles com sempre. Però el que queda àmpliament clar és que no som testimonis únicament d’una explosió de noms en un paper. L’impressionant registre fòssil de què disposem avui dia apunta realment a una diversitat d’espècies en la història dels homínids.
El gènere ‘Homo’ avui dia
Aquesta prova de diversitat ens porta de tornada a les preguntes centrals que aborda aquest article. Com es pot organitzar correctament la diversitat dels homínids? Quina part d’aquesta diversitat té cabuda de forma legítima en el nostre propi gènere Homo? I com podem reconèixer els límits d’aquest gènere? Com ja hem assenyalat, un gènere és una agrupació d’espècies relacionades per la seua ascendència. Però més enllà del requisit general de ser monofilètic, no hi ha un criteri general teòric ni pràctic que dicte com d’inclusiu hauria de ser un gènere. Llavors, on tracem la línia?
A pesar dels esforços d’investigadors com Bernard Wood i Mark Collard (vegeu Collard i Wood, 2015; Wood i Collard, 1999), que s’han pronunciat enèrgicament a favor de limitar el gènere Homo només a homínids que siguen adaptativament semblants a l’espècie tipus (Homo sapiens) i que es relacionen més estretament amb aquesta que no amb el tipus de qualsevol altre gènere, recentment es tendeix a la inclusió. De fet, l’ecumènic gènere Homo ja s’ha estès fins fa 2,8 milions d’anys (Villmoare et al., 2015). Però fins i tot suposant que la monofília siga rellevant en aquest cas, si volem mantindre qualsevol valor com un repositori d’informació filogenètica o estructural, el gènere Homo resultant, ampli i inclusiu, requeriria un elaborat aparell de classificació subgenèrica. El que, per descomptat, malbarata qualsevol intent d’agrupar tots els homínids relacionats. Per tant, em sembla molt més pràctic –a més de més defensable en termes teòrics– començar amb Homo sapiens i intentar conèixer quines altres espècies d’homínids seria útil classificar en un gènere que està necessàriament definit per la nostra pròpia espècie.
«Més enllà del requisit general de ser monofilètic, no hi ha un criteri general que dicte com d’inclusiu hauria de ser un gènere»
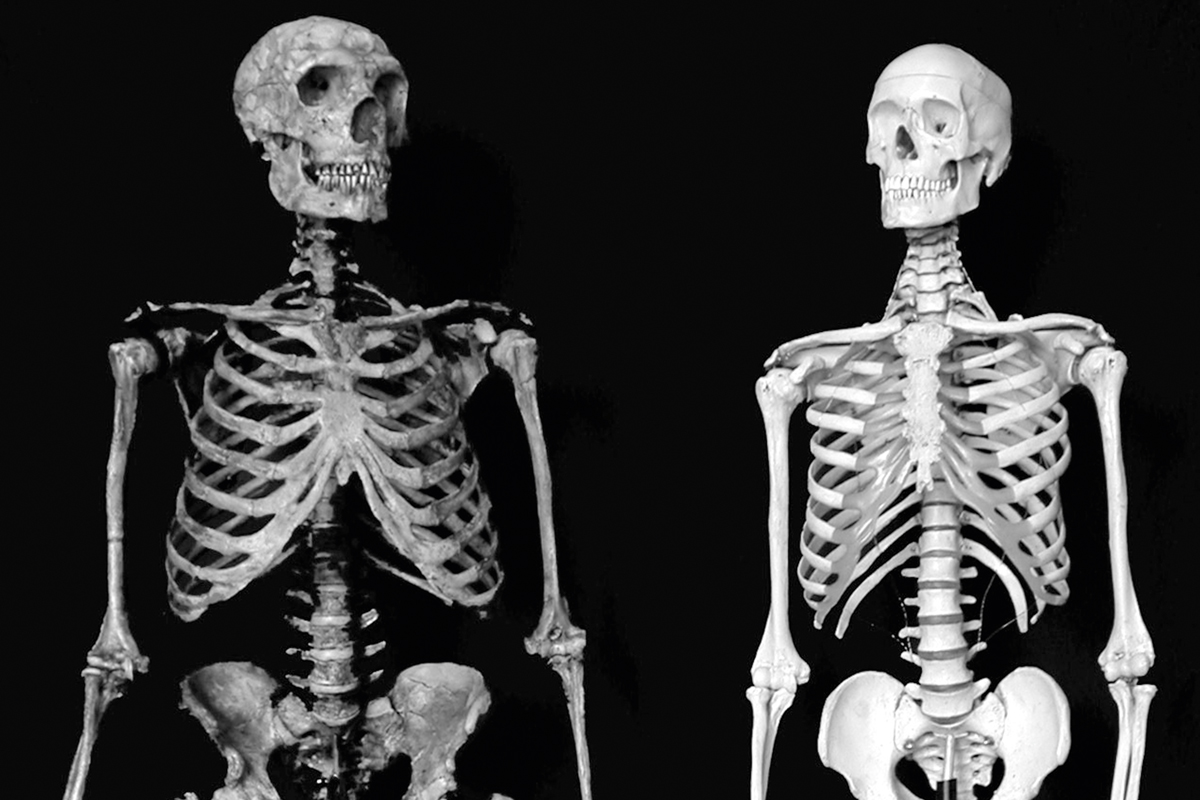
Inevitablement, el nombre d’espècies que caldria reconèixer entre els fòssils d’homínids més pròxims a Homo sapiens és un tema molt debatut. Per exemple, és clar que, dins del grup que es coneixia amb poques paraules com «Homo sapiens arcaic», hi ha representada una varietat d’homínids –encara que molt pocs considerarien avui un problema incloure totes aquestes formes en el gènere Homo–. El mateix ocorre amb els peculiars neandertals, avui dia reconeguts de manera quasi universal com a Homo neanderthalensis. Aquests homínids europeus de gran cervell estan estretament relacionats amb Homo sapiens, encara que ara està clar que el seu clade es va separar del llinatge que porta fins a Homo sapiens fa almenys mig milió d’anys (Arsuaga et al., 2014). També val la pena mencionar en aquest context que l’estructura osteològica d’Homo sapiens sembla haver derivat molt: quelcom que és evident en la comparació de la Figura 8 d’un esquelet d’humà modern amb un compost de neandertal que té un esquelet postcranial més primitiu. Una vegada més, però, ningú discuteix actualment que qualsevol d’aquestes diferències siga necessàriament un obstacle per a la inclusió d’ambdós en el gènere Homo.
Si anem més enllà, el debat continua obert sobre què hauria de constituir Homo erectus (l’encarnació moderna del Pithecanthropus erectus de Dubois). D’una. banda, se sosté que aquesta espècie és un tàxon estès per tot el vell món, i que comprèn una gran varietat de fòssils d’homínids d’Àsia i Àfrica al llarg de quasi dos milions d’anys. Altres, incloent-me a mi, dirien que hauríem de considerar Homo erectus una forma asiàtica endèmica i terminal, i en aquest cas els homínids africans anteriors, sovint assignats a Homo erectus –però morfològicament molt diferents del tipus de Java–, haurien de constituir la seua pròpia espècie, Homo ergaster (Tattersall, 2015a). Però una altra vegada sembla que no hi haja una raó teòrica convincent per la qual no podem veure tots els membres del grup erectus/ergaster com a membres d’un gènere Homo filogenèticament coherent. Perquè, encara que alguns d’ells posseïen cervells que podien assolir una grandària moderada (com a màxim), tots semblen haver adquirit esquelets de manera raonablement moderna. De fet, si les anàlisis i troballes futures confirmen que els homínids estan units per una configuració postcranial comuna, potser el millor seria reconèixer el gènere Homo no sols mitjançant apomorfies cranials, sinó també postcranials. Això podria resultar una mica incòmode per als éssers humans moderns, que valoren el que ocorre dins dels seus caps per damunt de tot. Però és possible que, al final, siga l’única manera de reconèixer un gènere Homo coherent en termes filogenètics i estructurals.
«Si les troballes futures confirmen que els homínids estan units per una configuració postcranial comunca, potser el millor seria reconèixer el gènere ‘Homo’ no sols mitjançant apomorfies cranials, sinó també postcranial»
Encara així, en incloure aquesta diversitat de fòssils ja estem començant a forçar els límits del que hauria de ser un gènere morfològicament coherent; i si ampliem el grup erectus/ergaster per a comprendre les sorprenents troballes d’homínids de Dmanisi (Lordkipanidze et al., 2013), observem un gènere Homo al qual s’ha forçat per poder encabir-hi una varietat poc convincent de grandàries cerebrals i morfologies cranials. El mateix s’esdevé fins i tot de manera encara més clara amb «Homo» floresiensis d’Indonèsia –d’una antiguitat inferior als 100.000 anys (Sutikna et al., 2016)– i amb «Homo» naledi de Rising Star (Sud-àfrica), que ara creiem que data de fa entre 335.000 i 236.000 anys (Dirks et al., 2017). És més, fins i tot després d’excloure del gènere Homo els homínids de Dmanisi, Flores i Rising Star, continuem tenint el conjunt de fòssils africans de fa 1,6 milions d’anys o més, restes que en les últimes dècades s’han atribuït a Homo habilis, a H. rudolfensis o simplement a H. sapiens. Leakey et al. (2001) va assignar rudolfensis al seu nou gènere Kenyanthropus, mentre que Collard i Wood (2015) en van excloure (amb bon criteri) totes aquestes formes d’Homo. No obstant això, un aclaparador consens continua atribuint tots aquests fòssils africans antics al nostre gènere.
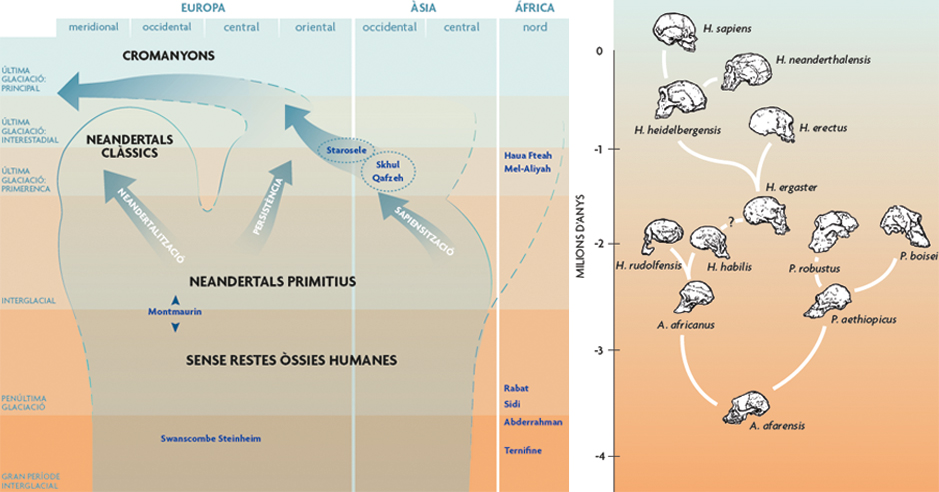 Figura 5. Diagrama de Clark Howell de 1957 sobre la distribució temporal i espacial d’alguns homínids del plistocè mitjà i superior. Noteu que Howell no afirma en cap moment que això siga un «arbre». En blau, jaciments amb conjunts de fòssils humans. Redibuixat per Kayla Younkin / Adaptat per Salvador SastreFigura 6. Filogènia molt temptativa de la família Hominidae, proposada per Ian Tattersall en 1993. / Dibuixat per Diana Salles / Adaptat per Salvador Sastre
Figura 5. Diagrama de Clark Howell de 1957 sobre la distribució temporal i espacial d’alguns homínids del plistocè mitjà i superior. Noteu que Howell no afirma en cap moment que això siga un «arbre». En blau, jaciments amb conjunts de fòssils humans. Redibuixat per Kayla Younkin / Adaptat per Salvador SastreFigura 6. Filogènia molt temptativa de la família Hominidae, proposada per Ian Tattersall en 1993. / Dibuixat per Diana Salles / Adaptat per Salvador Sastre
Quan, profundament influïts per la noció de «man the toolmaker» (“l’home creador d’eines”, vegeu Oakley, 1949), Louis Leakey i els seus col·legues van anomenar Homo habilis a la seua nova espècie de fa 1,8 milions d’anys, provinent de la gorja d’Olduvai a Tanzània (Leakey, Tobias i Napier, 1964), ho van fer seguint la tradició establida d’assignar formes fòssils al nostre gènere basant-se en comportaments inferits (dieta basada en carn, llenguatge, fabricació d’eines, art i simbolisme, etc.), més que no en la morfologia observable. No obstant això, la seua acció va desfermar un gran furor en una ciència que encara era captiva de Mayr. La majoria d’observadors o bé no volien separar aquests fòssils d’una espècie d’Homo erectus que el comprenguera tot, o bé rebutjaven reconèixer qualsevol distinció significativa respecte als espècimens d’Australopithecus de Sud-àfrica. Els partidaris d’aquesta segona opinió tenien un argument molt potent a favor seu, ja que sembla evident que la principal raó de Leakey i els seus col·legues per a assignar els fragments d’Olduvai a una nova, encara que més «primitiva», espècie d’Homo era que estaven suposadament associats amb les tosques eines de pedra trobades en la capa més baixa de la gorja.
Com que qualsevol nou comportament, fins i tot un fet tan radical com la fabricació d’eines de pedra, degué ser inventat necessàriament per un membre d’un gènere i una espècie existent (de qui no hauríem d’esperar diferències biològiques significatives respecte als seus pares o la seua descendència), el criteri de Leakey i els seus col·legues per a incloure una espècie en el gènere Homo mai va ser molt robust. No obstant això, per moltes raons (no gaire convincents), la noció d’Homo habilis va acabar quallant. A més, un variat assortiment de fòssils fragmentaris de fa, principalment, entre 2 i 2,5 milions d’anys es van atribuir al «primer Homo», especialment després que es descobriren les primeres eines de pedra en un lloc que data de fa més de 2,5 milions d’anys (Semaw et al., 1997).
«La dinàmica iniciada a Olduvai prompte va donar lloc a una enèrgica cerca del primer ‘Homo’ que continua encara avui»
La dinàmica iniciada a Olduvai prompte va donar lloc a una enèrgica cerca del «primer Homo» que continua encara avui. L’últim i més antic competidor per aquest lloc ha estat la forma Ledi-Geraru d’Etiòpia (Villmoare et al., 2015), amb una edat de 2,8 milions d’anys. No obstant això, cap dels pretendents a «primer Homo» té moltes raons morfològiques per a pertànyer a un gènere definit per l’altament derivatiu Homo sapiens. En canvi, sembla que, encara que els paleoantropòlegs no poden evitar reconèixer que hi ha diversitat en el registre dels primers homínids, continuen intentant dividir aquesta diversitat entre les dues categories establides: Australopithecus i Homo. I el desig d’encabir noves morfologies en aquestes dues caselles existents coste el que coste sembla més fort que l’evident necessitat d’ajustar el marc sistemàtic dels homínids (respecte a les espècies i als gèneres) per a donar-hi cabuda a noves morfologies.
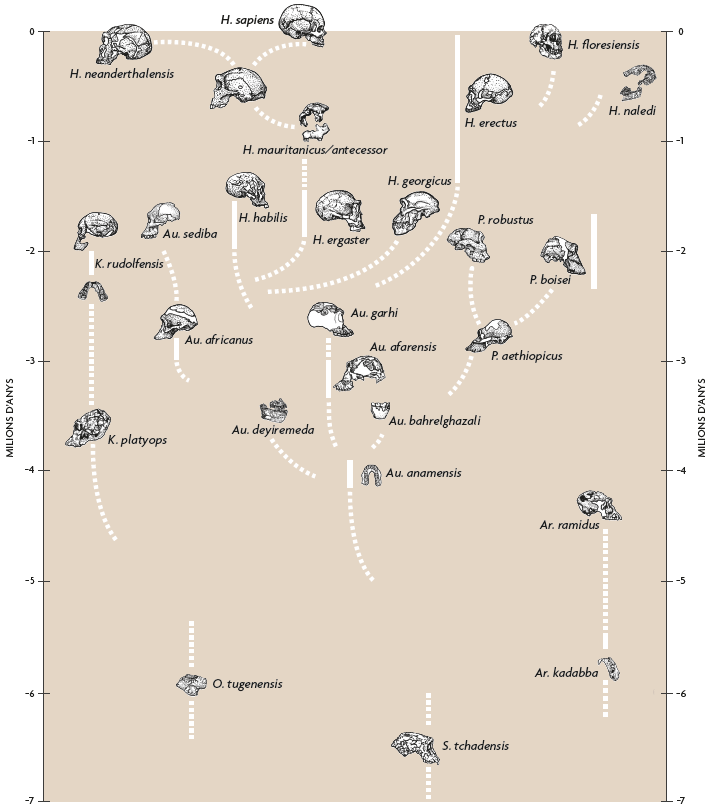
Figura 7. Filogènia molt temptativa de la família Hominidae, proposada per Ian Tattersall en 2017. Dibuixat per Kayla Younkin / Adaptat per Salvador Sastre
Així i tot, a pesar de la importància de la tradició, ara és clar que en el passat distant van existir múltiples llinatges d’homínids. I si volem entendre com s’estructurava aquesta diversitat, per descomptat serà millor analitzar-ho en termes de ramificació de clades, en compte d’aferrar-se de manera cada vegada més aparatosa a la tradició de la divisió entre els dos gèneres. Sí, en efecte, els gèneres són clades; però la diversitat morfològica que sorgeix del creixent registre fòssil homínid necessita clarament reflectir-se en un context sistemàtic més extens que el que permet la tradició (Schwartz i Tattersall, 2015; Tattersall 2015b, 2016). Al final, saturar un nombre limitat de gèneres canònics no ajudarà a aclarir l’evidentment complexa sèrie de successos que va portar a l’aparició d’Homo sapiens.
Conclusió
Al llarg de la història de la paleoantropologia, el gènere Homo s’ha contret i expandit, no al servei de la coherència filogenètica i morfològica, sinó seguint patrons filosòfics. Avui dia ens trobem en un extrem de l’expansió, amb un concepte absolutament ecumènic de gènere Homo que és, com a mínim, descurat, tant en termes filogenètics com morfològics. En un moment efímer va haver-hi senyals que començava una reavaluació temptativa del gènere i el seu contingut, amb la transferència d’alguns espècimens icònics d’Homo primitiu a gèneres com Australopithecus i Kenyanthropus (vegeu Leakey et al., 2001; Collard i Wood, 2015). Però des de llavors el pèndol s’ha encallat, i encara ens destorba un gènere que està literalment abarrotat, tant en l’aspecte morfològic com en el sistemàtic.
«Ara és clar que en el passat distant van existir múltiples llinatges d’homínids»
La lliçó que podem aprendre, o almenys hauríem de fer-ho, és que el fet de contindre Homo sapiens no converteix Homo en un gènere especial. Per tant, hem d’abandonar el nostre excepcionalisme homínid i definir el nostre propi gènere exactament igual com la resta. Aconseguir la coherència d’aquesta manera implicarà reduir el gènere Homo, centrant-se a reconèixer un tàxon fermament basat per un costat en la sinapomorfia i que, de l’altre, forma part d’un clade més complex que necessita més gèneres per a expressar la seua diversitat. Això, per descomptat, significa abandonar aquells candidats extravagants a «primer Homo» que només ho són perquè els seus fòssils no són d’australopitecins.




