

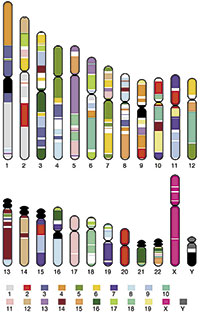
Figura 1. Comparació entre els cromosomes del ratolí i de l’home. Mitjançant el codi de colors, que corresponen als cromosomes del ratolí indicats en la part inferior, pot apreciar-se a quins fragments dels cromosomes humans són equivalents, entenent com a tal la conservació dels gens que s’hi contenen. Queda de manifest que des de la separació dels llinatges de rosegadores i humans s’han produït nombrosos canvis en la disposició cromosòmica, però el contingut total en gens d’ambdós organismes és molt semblant.
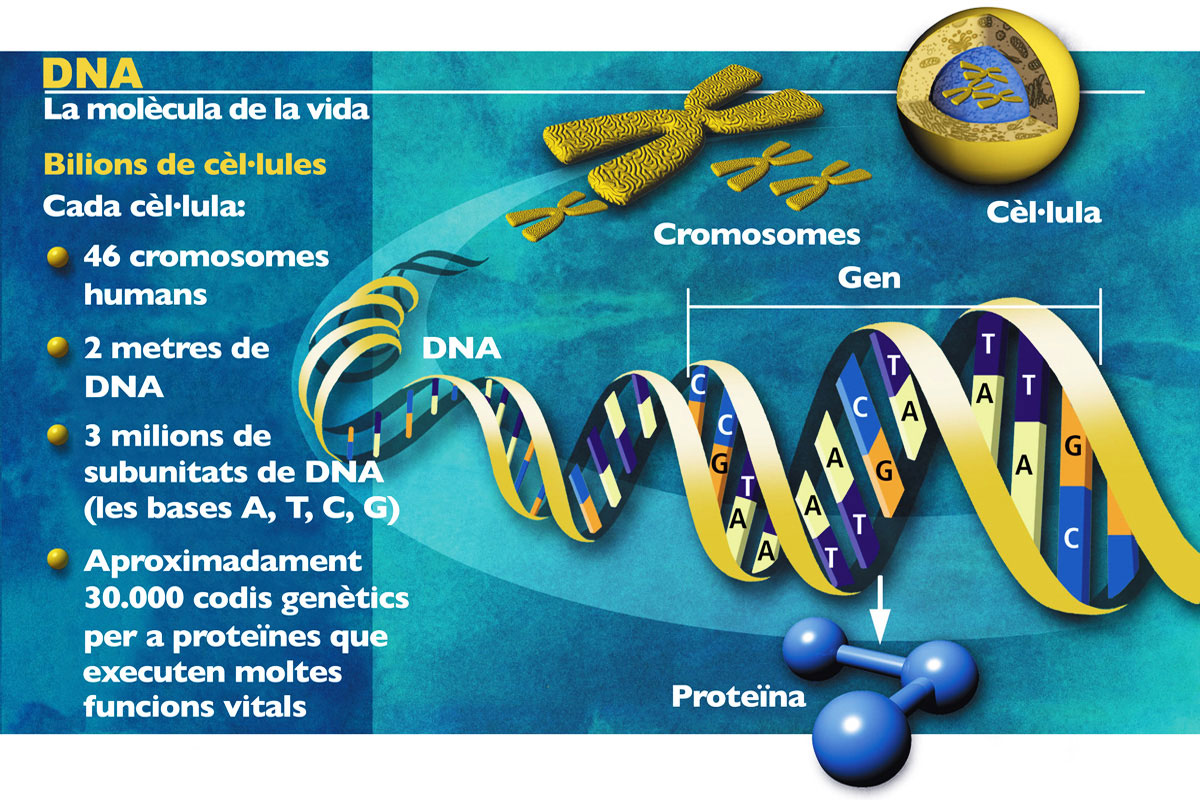
El genoma és el conjunt del material hereditari d’un organisme, la seqüència de nucleòtids que especifiquen les instruccions genètiques per al seu desenvolupament i funcionament i que són transmeses de generació en generació, de pares a fills. A més dels gens pròpiament dits, s’hi inclouen regions espaiadores, regions reguladores, restes de gens antany funcionals i moltes seqüències més de funció o paper encara desconegut, si és que en tenen cap. De fet, en el genoma humà, a penes l’1,5% del material hereditari té una funció codificant, és a dir, correspon al que solem entendre per gens. Per tant, el genoma d’un organisme és el dipositari de la informació que permet que cada organisme es desenvolupe i responga a les exigències imposades pel medi. Però, a més, el genoma és dipositari dels canvis que, al llarg de la història de l’espècie corresponent i de totes les seues antecessores, n’han permès la supervivència fins als nostres dies. En conseqüència, en el genoma s’emmagatzema informació de dos tipus: una d’immediata utilitat per a l’organisme i una altra que serveix com a registre històric d’aquest i dels seus ancestres (figura 1). Ambdós tipus d’informació són explotats per la biologia actual, tant al seu vessant funcional com en l’històric o evolutiu.
Al voltant del genoma es plantegen diverses qüestions que convé aclarir. La primera té a veure amb la seua capacitat per a determinar totalment o parcialment el funcionament de l’organisme. A manera d’analogia, podríem comparar el genoma amb els plànols d’una casa elaborats per un arquitecte en el seu estudi. El resultat final depèn de moltes decisions i intermediacions no sempre previsibles: la disponibilitat de materials en cada moment, la interpretació realitzada pel director d’obra, la solució adoptada davant d’algun imprevist, les modificacions introduïdes pel propietari, fins i tot les preferències estètiques d’aquests! Per tant, l’edifici final pot diferir de l’imaginat inicialment per l’arquitecte que el va planificar, però aquestes diferències es produeixen més fàcilment en detalls accessoris i menys en els fonamentals. Igualment, podem dir que el genoma d’un organisme conté un conjunt d’instruccions, però que la forma en què aquestes es porten a terme depèn al seu torn de contingències ambientals i històriques que poden portar a diferències entre plànols (o fragments d’ells) en principi iguals. En conseqüència, la naturalesa de les instruccions genètiques no és completament determinista en tots els casos, si bé hi ha una sèrie de processos en què sí que es compleix aquesta perfecta relació entre herència i expressió final.
Una altra qüestió que hem d’aclarir és que no hi ha una relació un a un entre gens i caràcters observables o, almenys, que aquesta relació dista molt de ser general. En algunes ocasions, un únic gen determina un caràcter completament: per exemple, el sistema sanguini ABO o el grup Rh són determinats per un sol gen, respectivament. Aquesta mateixa situació es presenta amb certes alteracions genètiques i amb el desenvolupament de patologies, la qual cosa facilita enormement el diagnòstic precoç i obre les possibilitats per a la teràpia genètica. Però molts caràcters, la gran majoria, incloent-hi moltes condicions d’interès per a la medicina o la psicologia, tenen una base poligènica, és a dir, no existeix “el gen” que determina el caràcter de forma unívoca, sinó que aquest és el resultat de l’acció simultània de molts gens, en ocasions centenars, no tots amb la mateixa participació i damunt s’hi ha d’afegir l’efecte de l’ambient adés comentat. Molts dels debats clàssics sobre el determinisme genètic dels trets de la conducta i la personalitat, o la intel·ligència, sorgeixen d’una apreciació incorrecta d’aquesta naturalesa dual de l’expressió dels caràcters.
«Del genoma humà, a penes l’1,5% del material hereditari té una funció codificant»
Finalment, i com a sistemes complexos que som tots els éssers vius, hem de considerar el paper que tenen les interaccions entre les fraccions components del genoma a l’hora d’especificar el resultat final, almenys en el seu component genètic. Sabem que un gen pot afectar més d’un caràcter i que un caràcter pot ser afectat per més d’un gen. Per tant, una modificació en un gen pot provocar alteracions en diversos caràcters, el que coneixem com a efectes pleiotròpics, i l’expressió d’un cert caràcter pot dependre de quines variants es troben presents en dos o més gens diferents, situació que dóna lloc al fenomen conegut com a epístasi. Donat el nombre de gens presents en qualsevol organisme (desenes de milers en el cas d’animals i plantes), s’obre un extraordinari ventall de possibilitats d’interacció entre dos o més gens. Actualment no hem arribat a assolir ni tan sols una idea aproximada del paper que exerceixen les epístasis en la gran majoria de caràcters fenotípics.
«No existeix “el gen” que determina el caràcter de forma unívoca, sinó que aquest és el resultat de l’acció simultània de molts gens»
Com s’estudien els genomes?
Si bé la seqüenciació del genoma humà marca una fita en la biologia, les contribucions d’altres disciplines científiques han estat imprescindibles per a aconseguir-la. El projecte s’ha beneficiat d’avenços en la química, la física, les matemàtiques, la informàtica, i ha donat lloc, fins i tot, al naixement d’una nova disciplina integradora, la bioinformàtica, sense la qual no s’hauria pogut culminar, com veurem posteriorment. No ens pot estranyar, doncs, que entre els integrants dels equips que han aconseguit completar la seqüència del genoma humà es troben científics de disciplines molt dispars. Això mateix es repeteix en els equips que han aconseguit desxifrar les seqüències d’altres organismes. L’estudi del genoma en la seua integritat és el resultat d’avenços tècnics i conceptuals que van començar fa uns vint-i-cinc anys, quan Fred Sanger i Walter Gilbert desenvoluparen dos mètodes per a obtenir la seqüència de nucleòtids d’un fragment petit de DNA. Al cap de poc, el primer d’ells va aconseguir seqüenciar el genoma d’un bacteriòfag, un virus que infecta cèl·lules bacterianes, el fX174. Immediatament van començar a acumular-se seqüències de genomes vírics, seguides, a principis dels anys 80 per la seqüència del genoma mitocondrial humà. L’obtenció de cada una d’aquestes seqüències representava la culminació d’una àrdua tasca de mesos, fins i tot anys, a pesar que aquests genomes només contenen uns pocs milers de nucleòtids. Atès que el genoma humà conté uns 3.000 milions de nucleòtids, el salt que representa passar de seqüenciar un genoma viral a l’humà semblava infranquejable el 1980, quan David Botstein i els seus col·laboradors van proposar aquest objectiu a la comunitat científica. L’interval de gairebé una dècada entre la proposta inicial i la decisió política final de portar-la a terme va permetre que es produïren avenços decisius en diversos fronts, com en les tècniques de mapatge físic, que permeten l’aïllament de gens i regions concretes a partir de la seua localització cromosòmica, i en les estratègies de seqüenciació mitjançant “perdigonades” aleatòries, que posteriorment van permetre d’automatizar un procés que fins llavors era manual.

Figura 2. Gràfic temporal que mostra els avenços en la seqüenciació de genomes complets. En l’actualitat el nombre de projectes de seqüenciació de genomes complets en marxa supera els dos centenars, als qual caldria afegir-ne molts més que porten a terme empreses privades i que no són fets públics a la comunitat.
A finals de 1990 es va llançar el Projecte Genoma Humà amb la creació de centres de seqüenciació als Estats Units, Regne Unit, França i Japó i amb el suport de la Comunitat Europea. En els cinc anys següents es va progressar ràpidament en dos fronts: d’una banda, la construcció de mapes genètics i físics dels genomes humà i de ratolí, la qual cosa va facilitar eines indispensables per a identificar gens lligats a malalties i va fixar fites per a la seqüenciació posterior. Un segon front va ser la seqüenciació de dos genomes d’organismes eucariotes, el llevat Saccharomyces cerevisiae, utilitzat per a obtenir el pa i la cervesa, entre altres aliments, i el cuc nematode Caenorhabditis elegans (figura 2). En aquell moment, l’estratègia favorita per a la seqüenciació del genoma humà es desenvolupava en dues fases (figura 3). En la primera (figura 4), el genoma es dividia en fragments de grandària adequada (de només uns centenars de milers de bases) que eren, al seu torn, seqüenciats mitjançant l’estratègia de la perdigonada. Per a fragments d’aquesta grandària, l’estratègia de la perdigonada necessita que cada nucleòtid siga seqüenciat unes quantes vegades, quantificades amb el factor de cobertura o redundància, amb l’objectiu que no queden regions del fragment sense seqüenciar almenys una vegada. A mesura que el fragment és major en grandària, hi ha una tendència a no augmentar la redundància tant com és necessari a fi de garantir que cap nucleòtid es quede sense seqüenciar, per la qual cosa hi haurà alguns buits en la seua seqüència final. D’igual manera, per a obtenir aquests fragments d’alguns centenars de milers de bases, se seguia una estratègia semblant, per la qual cosa cada regió genòmica havia d’estar representada diverses vegades si es volia que no se’n deixara cap sense analitzar. La segona fase completava l’estratègia cega de la perdigonada buscant de completar aquells possibles buits faltants en els dos nivells esmentats. Aquesta estratègia va ser l’adoptada pel consorci públic i havia de produir una primera seqüència cap al 2003.
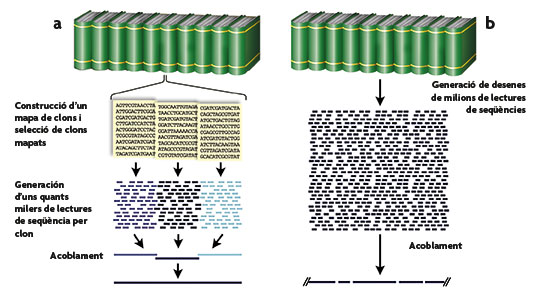
Figura 3. Comparació entre les dues estratègies de seqüenciació del genoma humà, la seguida pel consorci públic (a) i la del grup liderat per Craig Venter (b). En la primera, detallada amb més precisió en la figura 4, es parteix d’una col·lecció ordenada, mitjançant mapatge físic i genètic, de fragments de grandària mitjana, que són seqüenciats pel procediment de la perdigonada. Al contrari, en la segona estratègia es procedeix a la seqüenciació completa del genoma pel procediment de perdigonada, amb un nivell elevat de redundància per garantir la lectura de totes les posicions. De la tasca d’acoblar les seqüències fragmentàries obtingudes en una única seqüència lineal, se n’encarregaran poderosos sistemes informàtics. En ambdues estratègies es plantegen problemes amb les zones del genoma riques en DNA repetitiu, si bé en la primera és més fàcil identificar i quantificar l’extensió de les zones no seqüenciades per aquest motiu.
En 1998, Craig Venter, prèviament un dels líders del consorci públic i llavors ja en el sector privat, va llançar el desafiament que va conduir finalment a l’acceleració de tot el procés. La seua proposta consistia a abandonar les fases de mapatge físic i ordenació dels clons amb fragments grans per a passar directament a la seqüenciació completa del genoma mitjançant el mètode de perdigonada, deixant que nous algoritmes i ordinadors més potents s’encarregaren de l’acoblament de tota la seqüència. Rebuda amb escepticisme, almenys, la seua estratègia va demostrar la capacitat de seqüenciar un genoma complex en un temps rècord: va aconseguir seqüenciar el genoma (180 milions de nucleòtids) de Drosophila melanogaster, la mosca del vinagre i organisme model dels genètics des de principis del segle XX, en a penes un any. El seu grup va començar la seqüenciació del genoma humà (de fet, el genoma de cinc individus diferents) el 8 de setembre de 1999 i va concloure la fase d’obtenció de dades el 17 de juny del 2000. L’acoblament sobre el qual es va basar la publicació, simultània al resultat del consorci públic el 15 de febrer del 2001, es va completar en tres mesos i mig.
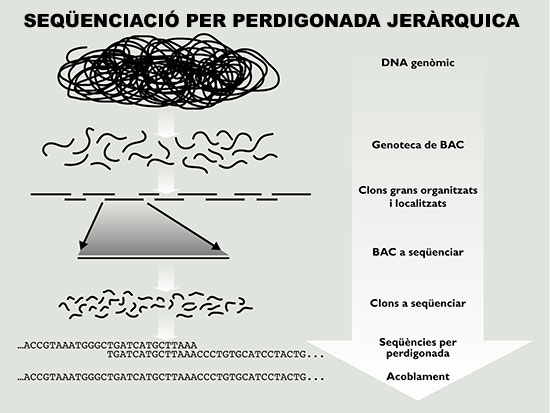
Figura 4. Esquema de l’estratègia de seqüenciació per perdigonada jeràrquica, la que ha aplicat el consorci públic. Es construeix una genoteca fragmentant el genoma (o cromosoma) desitjat i clonant-lo en vectors que accepten insercions grans (en aquest cas cromosomes bacterians artificials o BAC). Els fragments de DNA genòmic presents en la genoteca s’organitzen en un mapa físic i a continuació s’elegeixen els clons BAC individuals per seqüenciar-los per perdigonada. Finalment, les seqüències dels clons s’acoblen per a reconstruir la seqüència del genoma.
Al llarg del gairebé any complet que ha transcorregut des de la publicació, han aparegut nombrosos estudis basats en l’anàlisi comparativa d’ambdues seqüències del genoma humà, al mateix temps que, especialment entre alguns investigadors pertanyents al consorci públic, s’han suscitat seriosos dubtes sobre la factibilitat de la seqüència de la perdigonada per a un genoma tan gran com l’humà si el grup liderat per C. Venter no haguera tingut accés a la informació publicada pel consorci públic. Entre els estudis comparatius, sorprèn que, a pesar de la bona coincidència respecte del nombre de gens previstos per ambdós grups (entorn dels 35.000), bona part d’ells no coincideixen entre les dues prediccions. D’altra banda, algunes afirmacions, com l’existència d’uns 150 gens en el genoma humà més pròxims a gens de procariotes, bacteris en aquest cas, que a cap altre organisme evolutivament més emparentat amb nosaltres han estat clarament refutades.
Tot i que hem après molt i molt ràpidament, encara queden moltes incògnites per resoldre, moltes sorpreses amb què replantejar-nos la nostra concepció actual i molts detalls per revelar abans de poder utilitzar profitosament la immensa quantitat d’informació que tanca el genoma. Solament llavors estarem en condicions de comprendre, a tots els nivells, com funciona un ésser viu.
Les revistes Nature i Science han dipositat gran quantitat d’informació lliurement accessible en els seus servidors d’Internet.




