Sobreviure a la incertesa
Biodiversitat, adaptació i fluctuació ambiental en rotífers

L’estudi de l’evolució enfront de la incertesa ambiental és crucial per a entendre la diversitat biològica, ja que diversificar les estratègies de vida és clau per a sobreviure i reproduir-se en un ambient incert. Els rotífers són microinvertebrats planctònics que habiten masses d’aigua continentals. El seu cicle vital complex, que combina la reproducció sexual i l’asexual, juntament amb una grandària petita i temps de generació curts, els converteix en excel·lents organismes model en estudis d’ecologia evolutiva. Ací presentem estudis poblacionals de camp i d’evolució experimental en el laboratori on vam mostrar que aquests organismes són capaços d’adaptar-se localment diversificant el seu cicle vital enfront de fluctuacions impredictibles en el seu ambient.
Paraules clau: plàncton, rotífers, cicles vitals complexos, ambients temporals, impredictibilitat ambiental.
«Si un organisme fora capaç d’adaptar-se a qualsevol circumstància present a la biosfera, seria l’únic ésser viu a la Terra»
Introducció
Si un organisme fora capaç d’adaptar-se a qualsevol circumstància present a la biosfera –és a dir, poguera subsistir als tròpics i als pols, a l’aigua i a l’atmosfera, a partir de qualsevol font biòtica o abiòtica d’energia– seria probablement l’únic ésser viu a la Terra i la diversitat d’organismes en aquesta seria mínima. Contràriament, en la pràctica, adaptació significa especialització, i implica millorar les aptituds en relació amb unes condicions determinades, sacrificar unes capacitats per unes altres. Per això, una «dimensió», però no l’única, per a explicar la biodiversitat és l’adaptació. L’adaptació es requereix per a la persistència d’una espècie en una localitat, i en conseqüència en un hàbitat o en una regió, i en la biosfera. El darwinisme ens ensenya que l’adaptació ocorre per selecció natural. Aquesta, com qualsevol altra força natural, actua com el que un aristotèlic diria «causa eficient»,1 des del passat i sense finalitat. És a dir, quan un ésser viu es troba davant un ambient que d’alguna manera és «genuïnament» nou, no cal esperar que hi estiga adaptat. En el millor dels casos, entrarà en un procés de selecció natural que acabe adaptant-lo a la novetat. Com que tots els ambients són canviants, això representa un repte crucial per a l’ajust entre els organismes i el seu ambient.
«Adaptació significa especialització, i implica millorar les aptituds en relació amb unes condicions determinades»
Per a adonar-se de la ubiqüitat dels canvis ambientals n’hi ha prou de pensar en els propis del clima, i reparar en el que guardem en els nostres armaris (roba d’estiu, d’hivern, d’entretemps…) o considerar els nostres dubtes sobre què portar quan eixim d’excursió. Aquests dos exemples il·lustren casos de «fluctuacions ambientals», en els quals existeix una recurrència freqüent en les condicions, i que contrasten amb les «novetats ambientals», com ara la que podria representar un refredament del clima en escala temporal geològica o la irrupció d’un nou patogen. Mantenint el símil: no guardem roba en l’armari en previsió d’una època glacial. En el cas de les fluctuacions, la recurrència es dona dins de l’escala temporal pròpia de la resposta evolutiva dels organismes, i pot actuar com una pressió selectiva per la qual evolucione l’adaptació no solament a un valor mitjà de les condicions ambientals, sinó a la variabilitat d’aquestes.
«Si l’adaptació a la incertesa ambiental és important en qualsevol circumstància, serà probablement crucial en el futur»
En aquest article tractem l’adaptació a la fluctuació ambiental, però no de qualsevol tipus. Els exemples de l’armari i l’excursió il·lustren dues maneres molt diferents de fluctuació. Algunes són, bé per la seua periodicitat, bé perquè hi ha indicis que les anticipen, predictibles per als organismes. Unes altres no ho són. Així i tot, la mitjana i la variància (aquesta última, la mesura estadística més comuna per a quantificar la variació) d’un factor que fluctua impredictiblement poden bastar perquè, per selecció natural, aparega una optimització en els trets de l’organisme (optimització en el sentit que maximitza l’eficàcia biològica, a vegades denominada aptitud). La teoria evolutiva així ho apunta, i els nostres estudis empírics, com veurem, també. Però abans, advertim d’una complicació de segon ordre: una de les novetats ambientals que cal esperar com a conseqüència del canvi global és l’increment de les fluctuacions i la seua impredictibilitat. Si l’adaptació a la incertesa ambiental és important en qualsevol circumstància, serà probablement crucial en el futur.
Estratègies de minimització de riscos
Per a un organisme que necessàriament compleix el seu cicle vital en un any, de res li serveix deixar molts descendents la major part dels anys si uns altres no pot deixar-ne cap. Aquests annus horribilis representarien per a ell una barrera infranquejable. La seua eficàcia biològica, considerant el conjunt d’anys, seria zero. Per tant, la mitjana aritmètica de l’eficàcia biològica en les diferents situacions ambientals, sovint utilitzada per a descriure l’èxit reproductiu individual, no és en aquest cas una mesura adequada. En ambients impredictibles, esperem adaptacions que facen que els individus no maximitzen la seua eficàcia mitjana; les prediccions evolutives no poden fer-se sense tenir en compte la variància en l’eficàcia. Així, quan hi ha una necessitat d’anticipar-se a canvis impredictibles, s’espera l’evolució d’estratègies de minimització de riscos (bet hedging) que redueixen la variància de l’eficàcia a costa de disminuir la mitjana aritmètica de l’eficàcia, però que augmenten la seua mitjana geomètrica; és a dir, maximitzen l’eficàcia biològica a llarg termini.
«Per l’escala temporal a la qual opera, la resposta adaptativa dels organismes no és fàcil d’estudiar»
La teoria evolutiva ha investigat aquest problema amb diverses aproximacions. Una de ben simple, però que malgrat això copsa l’essència del problema, suposa que un organisme pot enfrontar-se a dos tipus de condicions (favorables o adverses), les quals se succeeixen amb certa probabilitat en una sèrie impredictible. Una manera de minimitzar els riscos és posseir unes característiques constants que funcionen bé qualssevol que siguen les condicions. Aquestes característiques permetrien a l’espècie persistir en les condicions adverses a costa de no explotar completament les condicions favorables seguint una estratègia conservadora. Una altra manera de minimitzar riscos és produir certa fracció de descendents ajustada a les condicions adverses i certa fracció ajustada a les favorables. Es tracta d’augmentar les opcions d’èxit amb una estratègia diversificadora. Així, Cohen (1966) va analitzar el problema d’una planta anual que passa l’hivern en forma de llavor, la qual germina en acostar-se la primavera sense informació sobre si la qualitat de l’any permetrà assolir la fase de floració i fructificació. La seua predicció va ser que en aquestes plantes una fracció de les llavors produïdes per l’adult hauria de germinar (per si l’any és bo), mentre que una altra fracció hauria de mantenir-se en latència (per si l’any és dolent), i aquest esquema de fraccionament hauria de repetir-se any rere any per a les llavors que han sobreviscut en latència l’any anterior. L’existència de bancs de llavors latents en els sòls és un indici que dona suport a aquesta predicció.
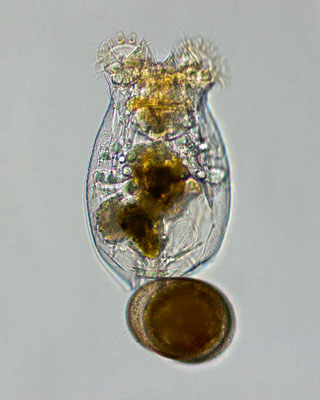
Figura 1. Microfotografia d’una femella de rotífer amb un ou de resistència. Aquests diminuts invertebrats solen formar part del plàncton de llacs i llacunes, i el seu cicle vital complex els converteix en excel·lents organismes model en estudis d’ecologia evolutiva. / Ivana Jezkova
Els rotífers, organismes model en ecologia i evolució
Per l’escala temporal a la qual opera, la resposta adaptativa dels organismes no és fàcil d’estudiar. O es recorre a inferències indirectes o es requereixen sistemes experimentals especials. Un d’aquests sistemes el proporcionen els rotífers (Serra, García-Roger, Ortells i Carmona, 2019), uns petits invertebrats (0,1-0,4 mm de longitud; Figura 1) que amb freqüència formen part del plàncton de llacs i estanys. La seua característica més interessant és la partenogènesi cíclica, el cicle vital de molts d’ells. La partenogènesi cíclica (Quadre 1) consisteix en la combinació de la reproducció sexual i l’asexual. Aquesta última, també anomenada partenogènesi –literalment, reproducció per verges2–, es basa en la producció de femelles que, excepte mutació, són còpies genètiques idèntiques de les seues mares. Sense necessitat d’aparellament i inversió en mascles, la reproducció asexual, en combinar-se amb la grandària corporal petita d’aquests organismes, dona taxes de proliferació elevades (la població pot duplicar-se en un parell de dies), que permeten una colonització ràpida de l’hàbitat i grandàries poblacionals nombroses. Això és crític pel fet que l’hàbitat dels rotífers és utilitzable tan sols durant un període del cicle anual, l’anomenada «estació de creixement», després de la qual la població activa desapareix. Abans que arribe el final de l’estació de creixement ha d’ocórrer almenys una generació sexual, la qual cosa en moltes espècies s’indueix per amuntegament a una determinada densitat poblacional. Llavors, naixen mascles que copulen amb femelles, els ous de les quals entren en latència i resisteixen en el sediment de la llacuna en condicions en les quals la supervivència d’individus actius és impossible. Comença llavors l’estació de latència, que pot ser prolongada. El moment de la reproducció sexual és, doncs, crític, prou tardà per a no comprometre la proliferació asexual i prou primerenc per a poder formar els ous resistents a temps. És clar que no hi ha dos anys iguals quant a la duració de l’estació de creixement. Aquesta duració és incerta.
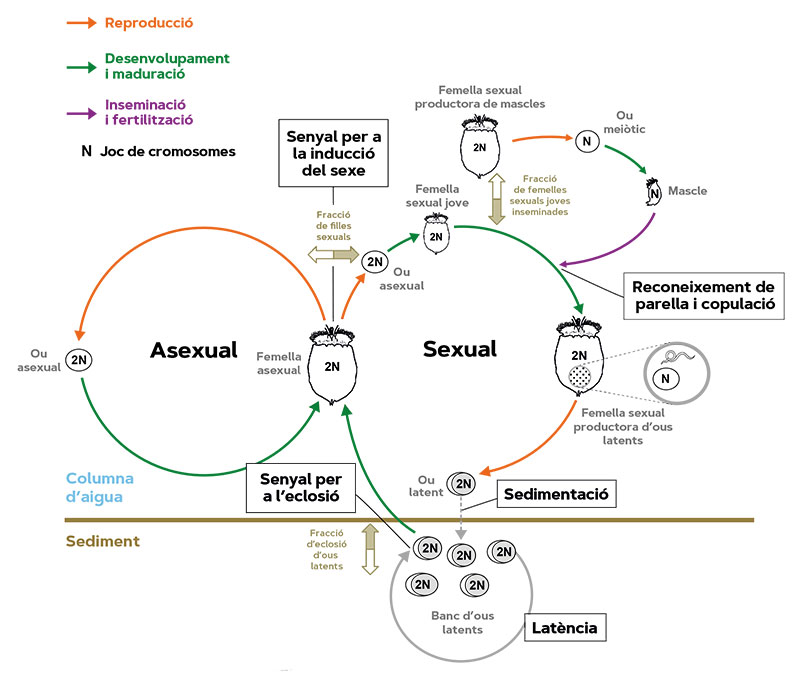
Quadre 1. Les poblacions dels rotífers partenogenètics cíclics són típicament temporals en el plàncton i recolonitzen la columna d’aigua durant l’estació de creixement planctònic mitjançant l’eclosió d’ous latents dels sediments de les llacunes i els estanys. Els nounats són femelles asexuals que produeixen ous diploides (amb dos jocs cromosòmics) que –excepte mutacions– es desenvolupen en filles genèticament idèntiques a les seues mares. Aquesta reproducció exclusivament asexual es repeteix un nombre indeterminat de vegades (proliferació clonal). La fase sexual –que no deté completament la reproducció asexual– comença amb la producció partenogenètica de filles sexuals per part de mares asexuals com una fracció de la seua descendència en resposta a factors ambientals inductors. Les femelles sexuals produeixen ous amb un únic joc de cromosomes (haploides) resultant de la meiosi (divisió cel·lular en la qual la dotació genètica es redueix a la meitat), que es desenvolupen partenogenèticament en mascles haploides o, si les femelles sexuals joves s’aparellen, els seus ous haploides són fertilitzats i es converteixen en ous latents diploides. Aquests ous es depositen en el sediment, poden sobreviure en condicions adverses i permeten a les poblacions recolonitzar la columna d’aigua quan es reprenen les condicions adequades en l’hàbitat i també dispersar-se a altres hàbitats. Després de rebre els estímuls apropiats, una fracció dels ous latents fa eclosió en femelles asexuals, i comença una nova estació de creixement. La fracció dels ous en latència que no fa eclosió s’acumula i forma bancs d’ous en el sediment. / Font: Modificat a partir de Serra et al. (2019).
«El cicle vital de molts rotífers és la partenogènesi cíclica, que consisteix en la combinació de la reproducció sexual i l’asexual»
La partenogènesi cíclica representa un delicat ajust entre els avantatges i desavantatges de la reproducció sexual i el seu tret concomitant en aquests animals: la latència. Aquest cicle vital mereix per si mateix estudi i s’ha utilitzat per a la investigació d’hipòtesis evolutives sobre els avantatges de la reproducció sexual, hipòtesi sobre la divergència entre poblacions locals, i hipòtesis ecològiques sobre la coexistència d’espècies (Serra et al., 2019). A més, el cicle vital dels rotífers ofereix l’oportunitat per a distingir fàcilment entre factors genètics i ambientals, ja que la reproducció asexual genera clons d’individus genèticament idèntics. Finalment, les altes taxes de proliferació asexual impliquen temps de generació curts. Això, al costat de la grandària tan petita dels rotífers, permet estudis d’evolució experimental; és a dir, estudis per a comprovar si poblacions de laboratori responen a pressions de selecció que imiten les que es troben en el seu hàbitat, però que són creades per l’investigador en condicions controlades.
Estudis amb poblacions ibèriques de rotífers

Figura 2. En la imatge, mostreig en la llacuna manxega d’Hoya Rasa, inclosa en els estudis del Laboratori d’Ecologia Evolutiva de l’Institut Cavanilles de Biodiversitat i Biologia Evolutiva de la Universitat de València. El mostreig de les poblacions actives de rotífers en la columna d’aigua es realitza amb una xarxa de zooplàncton (en la imatge, al fons) i el de les seues formes latents (ous de diapausa), en el sediment, amb una draga Van Veen. En la mateixa imatge, davant de tot, un investigador recollint el sediment extret amb la draga. / María José Carmona
El clima de la regió mediterrània es caracteritza per variacions irregulars que afecten el règim de precipitacions i evaporació. No és sorprenent que aquesta variació afecte la hidrologia de petites masses d’aigua com les llacunes de la Manxa (Figura 2), en l’est de la península Ibèrica. Per a l’estudi d’aquesta regió, Lluís Franch-Gras i col·laboradors (Franch-Gras, García-Roger, Franch, Carmona i Serra, 2017), membres del Laboratori d’Ecologia Evolutiva de l’Institut Cavanilles de Biodiversitat i Biologia Evolutiva (ICBiBE) de la Universitat de València, van adaptar un conjunt de mètodes d’anàlisi d’imatges de satèl·lit. L’objectiu era la detecció de masses d’aigua que són d’àrea variable. Això els va permetre aprofitar una sèrie temporal de dades inusualment llarga en ecologia. Mitjançant aquesta anàlisi, es van caracteritzar les irregularitats dels períodes anuals d’inundació (l’hidroperíode) de les llacunes, la qual cosa va revelar l’existència d’un rang ampli d’impredictibilitat, des de llacunes molt predictibles fins unes altres molt poc predictibles.
«Els genotips procedents de llacunes amb impredictibilitat ambiental iniciaven la reproducció sexual més primerencament que els procedents d’ambients més predictibles»
A partir d’aquestes observacions, va poder precisar-se (Franch-Gras, García-Roger, Serra i Carmona, 2017) una primera hipòtesi adaptativa, segons la qual les poblacions de rotífers que viuen en llacunes molt impredictibles tenen un inici de la reproducció sexual primerenc, com una manera de produir ous latents també primerencament i encarar amb èxit una estació de creixement inesperadament curta. Això, però, a costa de disminuir la reproducció asexual, i per tant d’explotar més plenament una estació de creixement potencialment llarga. En una segona hipòtesi, molt pròxima a la ja comentada per a plantes anuals, aquests investigadors van proposar que, en llacunes d’hidroperíode impredictible, els rotífers posseirien gens que els permetrien produir una fracció alta d’ous latents que no farien eclosió la primera vegada que les condicions del mitjà semblaren favorables. Així, s’estalviarien ous que romandrien latents en el sediment per si l’estació de creixement anara massa curta per a completar el cicle reproductiu amb la producció d’una nova cohort d’ous latents, això fins i tot a costa de perdre capacitats competitives si l’estació de creixement sí que permetera completar el cicle. Aquestes prediccions impliquen una diferenciació genètica entre poblacions d’una mateixa espècie, la qual cosa significa que es dona una adaptació local. La comprovació d’aquestes hipòtesis, per tant, ha de fer-se preferentment mantenint els rotífers procedents de les diferents llacunes en condicions de laboratori, controlant els efectes ambientals no selectius, però que afecten el desenvolupament dels organismes.
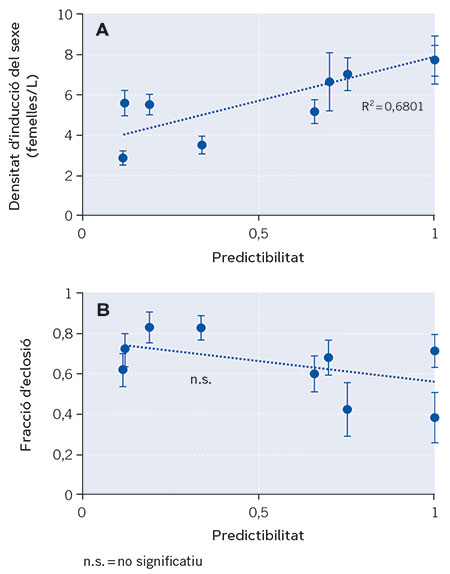
Figura 3. Relació entre dos trets relacionats amb la latència de nou poblacions naturals del rotífer partenogenètic cíclic Brachionus plicatilis i el grau de predictibilitat ambiental de les llacunes que habiten. En la figura A, inici de la reproducció sexual, inferit a partir dels valors de densitat poblacional als quals s’indueix. Un menor valor de densitat a la qual s’indueix la reproducció sexual indica un inici més primerenc d’aquest tipus de reproducció que garanteix la producció d’ous latents en aquests rotífers. En la figura B, pot observar-se la fracció d’eclosió d’ous latents (en tant per u). Una fracció menor garanteix la supervivència si l’ambient és desfavorable. De cada tret es mostren els valors mitjans per a cada població i les seues barres d’error (±1 error estàndard). La línia discontínua en cada gràfica representa l’ajust per regressió minimoquadràtica. El coeficient de determinació (R2 = 0,6801) és la proporció de la variància en la densitat d’inducció del sexe explicada per la predictibilitat ambiental. / Font: Modificat a partir de Franch-Gras et al. (2019).
Els resultats van mostrar que, efectivament, els genotips procedents de llacunes amb impredictibilitat ambiental iniciaven la reproducció sexual més primerencament que els procedents d’ambients més predictibles (Figura 3a). No obstant això, no es va obtenir evidència a favor de la segona hipòtesi (Figura 3b), potser perquè n’hi ha prou amb un tret –el moment d’inici de la reproducció sexual– per a assolir l’estratègia de minimització de riscos necessària, o potser perquè l’heterogeneïtat de condicions en el sediment és tan elevada que un tret intrínsec –com és la fracció d’eclosió– és poc eficaç (Franch-Gras, García-Roger, Serra, et al. 2017).
L’evidència proporcionada pels estudis realitzats al nostre laboratori (Franch-Gras, García-Roger, Franch et al., 2017; Franch-Gras, García-Roger, Serra et al., 2017) és observacional, la qual cosa implica que les relacions es descriuen en el seu context natural –són realistes–, però cal preguntar-se si aquestes relacions podrien no ser causals; és a dir, que els trets del cicle vital s’hagueren diferenciat entre poblacions per alguna altra pressió selectiva concomitant amb la impredictibilitat ambiental. Per a descartar això es van dur a terme estudis d’evolució experimental (Tarazona, García-Roger i Carmona, 2017). En el laboratori, es van crear ambients amb fluctuació aleatòria en la longitud de l’hidroperíode i ambients amb una longitud de l’hidroperíode constant. Les poblacions de rotífers hi van créixer en una sèrie de cicles d’inundació. La biologia dels rotífers va permetre simular aquesta complexitat en quimiòstats mantinguts en una cambra amb condicions controlades. Els resultats van corroborar les prediccions i a més van mostrar que la divergència adaptativa esperada va ser ràpida. Les poblacions de laboratori van evolucionar cap a un inici de la reproducció sexual més primerenc en condicions impredictibles, d’acord amb l’estratègia de minimització de riscos conservativa apuntada pels resultats de l’estudi en poblacions naturals (Figura 4a). A més, les poblacions en el règim impredictible també van evolucionar cap a fraccions d’eclosió d’ous latents menors que les del règim predictible (Figura 4b). En altres estudis s’ha reconegut la rapidesa d’evolució dels rotífers (Declerck i Papakostas, 2016). La reproducció sexual combinada amb una selecció clonal que opera sobretot el conjunt dels gens d’un individu són factors importants. En el nostre sistema influeix sens dubte el que la població experimental inicial estava composta de diversos clons procedents de diverses poblacions naturals. A diferència de les observacions en el camp, en els estudis d’evolució experimental es van complir les dues prediccions formulades. Probablement, les condicions de laboratori en les quals fan eclosió els ous latents, molt més homogènies que les de la naturalesa, fan que el genotip s’expresse més netament, i la fracció d’eclosió siga un bon blanc per a l’evolució per selecció.
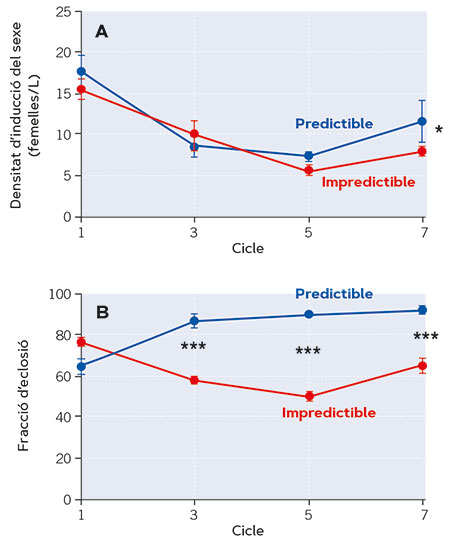
Figura 4. Evolució de dos trets relacionats amb la latència en poblacions de laboratori de Brachionus plicatilis al llarg de set cicles de selecció en règims divergents de fluctuació ambiental en relació amb la duració de l’hidroperíode: predictible (duració constant de l’hidroperíode al llarg dels cicles de creixement) i impredictible (hidroperíode de duració variable al llarg dels cicles de creixement). En la Figura A pot observar-se l’inici de la reproducció sexual, inferida a partir dels valors de densitat poblacional al llarg dels cicles d’evolució experimental. Un menor valor de densitat a la qual s’indueix la reproducció sexual indica un inici més primerenc d’aquesta. Així, en general, en condicions més impredictibles, l’inici de la reproducció sexual sol ser més precoç. En la Figura B, fracció d’eclosió d’ous latents (en percentatge) al llarg dels cicles d’evolució experimental. Pot observar-se que, en els cicles impredictibles, la fracció d’ous que fa eclosió és menor (és a dir, més quantitat es mantenen en latència per a mantenir-se a resguard de condicions potencialment desfavorables). De cada tret es mostren els valors mitjans per a cada règim i les seues barres d’error (± 1 error estàndard). Els asteriscos indiquen diferències estadísticament significatives entre règims (*p < 0,05; **p < 0,01; ***p < 0,001). / Font: Modificat a partir de Tarazona et al. (2017).
El disseny dels estudis esmentats (combinació de treball de camp i de laboratori) permet conèixer de manera no ambigua que les diferències en els trets de cicle vital són genètiques, és a dir, que les diferències no es confonen per variacions en l’ambient en què s’expressen els gens. Per la seua banda, els resultats ofereixen una evidència forta que aquestes diferències en el cicle vital són degudes a la selecció natural en resposta a la impredictibilitat ambiental; és a dir, són degudes a una divergència evolutiva adaptativa. Així i tot, aquests estudis no van permetre identificar els gens específics responsables. En l’actualitat, el desenvolupament de les tecnologies d’anàlisi genòmica –en els seus dos vessants, la seqüenciació del material genètic i la seua anàlisi bioinformàtica– permet avançar en aquest últim sentit fins i tot per a espècies que no han estat objecte d’un estudi genètic intensiu.
Així, en un nou estudi (Franch-Gras et al., 2018) es va seqüenciar i es va elaborar un «esborrany» de genoma per a l’espècie estudiada i després es va analitzar la divergència entre els genomes de les poblacions prèviament caracteritzades per la seua adaptació diferencial a la impredictibilitat ambiental. Utilitzant genotipatge per seqüenciació i la subsegüent anàlisi bioinformàtica, es va trobar un gran nombre de posicions nucleotídiques en les quals hi havia divergència (polimorfismes de nucleòtid únic, o SNP en les seues sigles en anglès), fins i tot aplicant mètodes molt conservatius. Aquest procediment també va ser seguit per Tarazona i colaboradors per a les poblacions de laboratori que havien divergit en els assajos d’evolució experimental. En els dos estudis, diversos SNP –la majoria dels quals situats dins de gens– van mostrar una diferenciació entre poblacions major de l’esperada per atzar, i es van correlacionar amb la impredictibilitat ambiental i els trets del cicle vital estudiats. Per això, aquests gens són candidats a ser responsables de l’adaptació local a ambients impredictibles. Si bé la relació funcional d’aquests gens amb els trets del cicle vital no s’ha aclarit, representen un punt de partida per a estudis que ja cauen més en el camp de la biologia molecular i cel·lular que en el de l’ecologia evolutiva.
En el seu conjunt, els resultats mostren que les poblacions de rotífers poden desenvolupar respostes adaptatives en trets relacionats amb la latència per a ajustar-se localment al grau d’impredictibilitat ambiental. Aquestes respostes impliquen diferents tipus d’estratègies de minimització de riscos. Els estudis duts a terme en el laboratori d’Ecologia Evolutiva de l’ICBiBE proporcionen una anàlisi de la resposta evolutiva d’aquestes poblacions, utilitzant una aproximació integrada en què es cartografia el genotip (marcadors genètics) en el fenotip (trets del cicle vital), i el fenotip en l’ambient (trets de l’hàbitat).
«Hi ha forces evolutives que poden oposar-se a la selecció natural i dificultar l’adaptació»
Aparellament preferencial i protecció de l’adaptació local
Hi ha forces evolutives que poden oposar-se a la selecció natural i dificultar l’adaptació. Per a les poblacions de rotífers abans esmentades, i que habiten llacunes molt pròximes entre si, n’hi ha prou de pensar quin efecte tindria sobre una població (la resident) l’arribada d’individus procedents de llacunes amb diferent predictibilitat. Aquests individus no estarien adaptats a la seua nova destinació, però així i tot podrien tenir alguna oportunitat de, en arribat el moment de la reproducció sexual, encreuar-se amb els residents. El resultat d’aquesta exogàmia seria una difuminació de la divergència adaptativa, i una pèrdua d’eficàcia per a la població resident. Per a aquest escenari s’ha postulat l’anomenada «hipòtesi del reforç», molt important en la teoria de l’especiació (Butlin, 1987). En breu, el procés descrit representa un cert aïllament reproductiu, ja que la descendència exogàmica no està adaptada a la localitat. Això ocasionaria una pressió de selecció en contra que residents i immigrants arribaren a aparellar-se, la qual cosa reforçaria l’aïllament preexistent. Si l’aïllament s’aprofundeix prou, les poblacions divergents arribarien a constituir espècies diferents, i així es generaria diversitat biològica. En el Laboratori d’Ecologia Evolutiva de l’Institut Cavanilles, Ivana Jezkova desenvolupa la seua tesi doctoral comprovant si els rotífers de poblacions locals divergents mostren indicis d’aparellament preferencial intrapoblacional en raó de la impredictibilitat ambiental (Figura 5). Els seus resultats preliminars apunten en aquest sentit i, si es confirmen, mostrarien un mecanisme d’estabilització de la divergència adaptativa enfront de la «pertorbació» de l’adaptació que significa la immigració. Els resultats d’aquest tipus són tremendament interessants perquè la hipòtesi del reforç no és trivial. En contra seua hi ha el fet que la reproducció sexual exigeix la coordinació de mascles i femelles, la seua coincidència temporal i espacial, i la identificació de parella. Aquesta coordinació penalitza la variabilitat.

Figura 5. En el reconeixement de parella en els rotífers intervenen quimioreceptors, proteïnes en la superfície corporal que actuen com a senyals mitjançant els quals el mascle (de menor grandària que la femella) reconeix les femelles de la seua mateixa espècie. En la imatge, s’observa el comportament de festeig que inclou (d’esquerra a dreta) la trobada, girs al voltant de la femella (circling) i la còpula. / Ivana Jezkova
«La reproducció sexual exigeix la coordinació de mascles i femelles, la seua coincidència temporal i espacial, i la identificació de parella»
Conclusions
L’evolució per selecció natural, si bé deixa el registre de la seua acció en el material genètic dels organismes, té un dels seus aspectes més cridaners en l’adaptació. Aquesta només pot entendre’s completament observant els trets morfològics, fisiològics i etològics dels organismes, però, especialment, observant també característiques del cicle vital, atès que aquestes característiques estan directament relacionades amb l’eficàcia biològica (és a dir, aquestes característiques són el que es denomina «components d’eficàcia»). Lliure de tot finalisme, aquesta teoria explica l’adaptació per dues consistències. D’una banda, la consistència entre l’ambient passat, que va seleccionar determinats trets, i l’ambient present, en el qual es manifesta el valor adaptatiu d’aquests trets. D’altra banda, la consistència en les característiques biològiques en els llinatges, deguda a l’herència. Això converteix en més desafiador –i més fascinant d’estudiar– l’adaptació a la fluctuació ambiental, sobretot si aquesta és irregular. La teoria, amb la seua capacitat per a idealitzar processos, apunta al fet que aquesta adaptació és en principi possible. Els experiments d’evolució duts a terme pel Laboratori d’Ecologia Evolutiva mostren que es pot donar en éssers vius –com els rotífers– compromesos amb moltes necessitats adaptatives més. Al seu torn, els patrons descrits pels estudis de poblacions naturals apunten al fet que les causes identificades en els experiments actuen també en l’entramat de factors que ocorre en la naturalesa. Falta esclarir els mecanismes poblacionals d’estabilització i –en un altre nivell– els mecanismes moleculars i cel·lulars subjacents. Pel que fa a això últim, en l’era de les «-òmiques» (genòmica, proteòmica, transcriptòmica, metabolòmica…) s’obre una tremenda oportunitat per a segellar el hiat entre la biologia molecular i l’ecologia evolutiva.
«Les poblacions de rotífers poden desenvolupar respostes adaptatives per a ajustar-se localment al grau d’impredictibilitat ambiental»
Un missatge final és que, enfront de la incertesa ambiental, l’evolució pot produir diversitat a diferents nivells: intraespecífica, que afavoreix la diversificació de les estratègies de vida (diversitat funcional) i interespecífica, que afavoreix els processos d’especiació (és a dir, la diversitat clàssica d’espècies).
1. En la seua Metafísica, Aristòtil descriu quatre classes de respostes enfront del perquè de les coses. La causa material té a veure amb el material del qual estan fetes les coses (és a dir, la causa d’una taula és la fusta); la formal té a veure amb el seu disseny (és a dir, la forma, disposició o aparença; per exemple, formalment una taula és una estructura plana recolzada sobre el terra per potes); la causa eficient té a veure amb la interacció amb altres agents externs (en el cas de la taula, la causa eficient és el fuster), i la causa final té a veure amb la finalitat de les coses, la qual cosa invoca l’existència d’un dissenyador (és a dir, la causa formal és la intenció del fuster de fer una taula per a menjar o estudiar). (Torna al text)
2. Del grec parthenos (“verge”) i genesis (“creació”). (Torna al text)
Part del treball que es presenta es va finançar amb fons públics del Ministeri Espanyol d’Economia i Competitivitat (projectes CGL2012-30779 i CGL2015-65422-P, tots dos cofinançats amb fons FEDER).
Referències
Butlin, R. (1987). Speciation by reinforcement. Trends in Ecology & Evolution, 2(1), 8–13. doi: 10.1016/0169-5347(87)90193-5
Cohen, D. (1966). Optimizing reproduction in a randomly varying environment. Journal of Theoretical Biology, 12(1), 119–129. doi: 10.1016/0022-5193(66)90188-3
Declerck, S., & Papakostas, S. (2016). Monogonont rotifers as model systems for the study of microevolutionary adaptation and its eco-evolutionary implications. Hydrobiologia, 796(1), 131–144. doi: 10.1007/s10750-016-2782-y
Franch-Gras, L., García-Roger, E. M., Franch, B., Carmona, M. J., & Serra, M. (2017). Quantifying unpredictability: A multiple model approach for Mediterranean ponds by using satellite imagery. PLOS ONE, 12(11), e0187958. doi: 10.1371/journal.pone.0187958
Franch-Gras, L., García-Roger, E. M., Serra, M., & Carmona, M. J. (2017). Adaptation in response to environmental unpredictability. Proceedings of the Royal Society B: Biological Sciences, 284(1868), 20170427. doi: 10.1098/rspb.2017.0427
Franch-Gras, L., Hahn, C., García-Roger, E. M., Carmona, M. J., Serra, M., & Gómez, A. (2018). Genomic signatures of local adaptation to the degree of environmental predictability in rotifers. Scientific Reports, 8(1), 16051. doi: 10.1038/s41598-018-34188-y
Franch-Gras, L., Tarazona, E., García-Roger, E. M., Carmona, M. J., Gómez. A., & Serra, M. (2019). Rotifer adaptation to the unpredictability of the growing season. Hydrobiologia. doi: 10.1007/s10750-019-3886-y
Serra, M., García-Roger, E. M., Ortells, R., & Carmona, M. J. (2019). Cyclically parthenogenetic rotifers and the theories of population and evolutionary ecology. Limnetica, 38(1), 67–93. doi: 10.23818/limn.38.13
Tarazona, E., García-Roger, E. M., & Carmona M. J. (2017). Experimental evolution of bet hedging in rotifer diapause traits as a response to environmental unpredictability. Oikos, 126, 1162–1172. doi: 10.1111/oik.04186
Els quatre autors formen part del Laboratori d’Ecologia Evolutiva de l’ICBiBE, la investigació del qual se centra en estudis d’ecologia evolutiva i ecologia de poblacions de microorganismes aquàtics, en concret, en l’anàlisi demogràfica, genètica i ecològica del zooplàncton. Entre els seus interessos científics destaquen l’adaptació dels cicles vitals dels rotífers a ambients variables, els processos mediadors de la coexistència d’espècies competidores, la diferenciació poblacional i especiació, la latència com a estratègia de dispersió en l’espai i en el temps i els processos evolutius que mantenen la reproducció sexual en les poblacions.