

Plantes a la carta
L’edició de genomes per a la millora vegetal

Les plantes que mengem són el resultat d’un llarg procés de domesticació d’espècies silvestres realitzat pels éssers humans. En l’actualitat la tecnologia d’edició de gens CRISPR/Cas proporciona una nova aproximació per a la millora dels cultius alhora que ofereix un interessant espectre de possibilitats en l’obtenció de varietats amb característiques noves i més saludables. La tecnologia es recolza en dos pilars fonamentals: d’una banda, el coneixement de la seqüència de genomes complets i, d’una altra, la identificació de la funció dels gens. En menys d’una dècada, dissenyar plantes a la carta ha passat de ser una utopia a una possibilitat real.
Paraules clau: cultius, millora genètica, CRISPR/Cas9, edició de gens.
Les plantes que mengem
Les plantes són components essencials dels ecosistemes terrestres i imprescindibles per al manteniment de la vida en la Terra. El regne Plantae inclou una enorme diversitat de grups: briòfits (molses i hepàtiques), pteridòfits (falagueres i equisetòpsids), gimnospermes i l’extens grup de les plantes amb flors o angiospermes (monocotiledònies i dicotiledònies). La major diversitat d’espècies, i possiblement el major nombre d’espècies encara sense identificar, es concentra en les zones tropicals del planeta.
«Malgrat l’aparent varietat de plantes utilitzades, únicament se’n consumeixen prop de 200 a tot el món de manera habitual»
S’estima que hi ha entre 450.000 i 500.000 espècies diferents de plantes,1 més de la meitat de les quals són comestibles. Malgrat l’aparent varietat de plantes que s’utilitzen en l’alimentació humana, únicament entorn de 200 es consumeixen a tot el món de manera habitual. A més, tres plantes –dacsa, arròs i blat– aporten quasi un 60 % de les proteïnes i calories d’origen vegetal en la dieta de les persones (FAO, 1999). La pèrdua d’agrobiodiversitat vegetal és una tendència que ha anat en augment en els últims cent anys i que està associada als canvis produïts en les pràctiques de cultiu i a la globalització dels sistemes de producció. El resultat és el reemplaçament de múltiples varietats locals per un nombre reduït de noves varietats millorades o per varietats exòtiques.
Els reptes actuals de l’agricultura passen per erradicar la fam i la malnutrició i assegurar aliments suficients per a donar resposta a la demanda de les generacions presents i futures (FAO, 1983). En les estimacions de la inseguretat alimentària en el món elaborades per la FAO en 2012 s’aprecia que el creixement econòmic és particularment eficaç per a reduir la fam i la malnutrició. D’altra banda, el creixement econòmic ha d’incloure la dimensió de la nutrició i incidir en la diversificació de la dieta i en l’adequada ingesta de nutrients dins de sistemes de producció i consum sostenibles (FAO, FIDA i PMA, 2012).
En anys recents, la prevalença de les al·lèrgies alimentàries i els seus efectes globals en la població mundial han estat objecte d’anàlisi. Les sensibilitats a aliments es poden classificar com a al·lèrgies si impliquen reaccions immunològiques anormals a components de l’aliment o com a intoleràncies si l’individu presenta sensibilitat a aliments en absència de resposta immunològica. La sensibilitat a productes d’origen vegetal inclou alguns dels aliments bàsics en la dieta humana, com els cereals amb glútens (blat, ordi, sègol, civada, espelta i les seues varietats híbrides o subproductes), la soja i la fruita seca, com els cacauets i les nous. En particular la intolerància al gluten afecta l’1 % de la població europea. El tractament recomanat per a les persones al·lèrgiques o intolerants és l’eliminació de la dieta de l’aliment o additiu que provoca la reacció adversa. En aquest sentit, la indústria alimentària ha de proporcionar la major quantitat d’informació en l’etiquetatge dels seus productes (Taylor i Hefle, 2001).
Innovació en millora vegetal
La millora de les espècies vegetals per a consum és una pràctica ancestral que ha tingut com a resultat la generació de plantes que presenten caràcters nous (Figura 1). S’ha utilitzat per a obtenir plantes de més qualitat, amb més rendiment o més resistents a condicions ambientals adverses i a patògens. A més, la millora vegetal també s’ha dirigit a obtenir varietats de plantes amb caràcters totalment nous (color i morfologia de la planta o el fruit, absència de llavors, etc.), no presents en l’espècie original. Les tècniques convencionals de millora vegetal obtenen la seua major font de variabilitat genètica de les mutacions que es poden originar de manera espontània durant els processos de mitosi o meiosi o mitjançant mutagènesi provocada (utilitzant compostos químics o irradiació).
«La millora de les espècies vegetals és una pràctica ancestral que ha tingut com a resultat la generació de plantes amb caràcters nous»
Sovint la mutació associada al caràcter desitjat ha d’incorporar-se en una altra planta mitjançant hibridació i posterior selecció en la població resultant de l’encreuament. D’aquesta manera es transfereixen un o pocs gens des d’una varietat o espècie donant a una altra varietat que actua com a receptor (parental recurrent). Es requereixen diverses generacions en les quals es torna a creuar la descendència amb el parental recurrent i se selecciona la presència del caràcter que volem introduir. Aquest procés té limitacions quant a la precisió dels fragments del genoma introduïts en el recurrent i requereix llargs períodes de temps per a introduir el caràcter desitjat. En els últims anys, el desenvolupament dels mètodes de seqüenciació massiva ha permès conèixer els genomes complets de molts cultius. Aquesta informació permet combinar les metodologies de millora convencional amb eines moleculars i accelerar els processos de selecció en els materials generats.
L’aparició de les tècniques d’enginyeria genètica va representar un salt substancial en la capacitat per a generar noves varietats de plantes genèticament modificades o GMO (organismes genèticament modificats). Durant els últims trenta anys el treball de nombrosos laboratoris ha donat lloc al desenvolupament d’una gran varietat de plantes amb modificacions d’interès; no obstant això, encara que la seua implantació per a ús comercial ha estat molt important, es redueix a una dotzena de cultius diferents (Beltrán, 2018). Les dues principals línies de millora de cultius s’han centrat en l’obtenció de plantes tolerants a herbicides i plantes resistents a insectes.
Una de les principals causes del baix nombre de varietats modificades presents en el mercat és l’elevat cost dels processos d’obtenció de permisos per a l’autorització de noves varietats. D’altra banda, la percepció social, principalment a Europa, que els aliments transgènics poden significar un risc per a la salut n’ha frenat la utilització i l’aparició de noves varietats. No obstant això, una revisió exhaustiva d’estudis científics publicats durant aquests anys no ha trobat cap evidència que les varietats obtingudes mitjançant enginyeria genètica representen un risc per a la salut o per al medi ambient major que les varietats obtingudes per millora convencional (National Academies of Sciences & Medicine, 2016).
L’edició de gens en plantes de cultiu: innovació i millora
Des de fa escassament sis anys, una nova tecnologia permet l’edició precisa de gens. La tecnologia CRISPR/Cas2 (CRISPR-associated) fa possible la millora de varietats vegetals sense la introducció d’elements forans. En particular, el sistema CRISPR/Cas9, per la seua simplicitat (Figura 2), baix cost i aplicabilitat a un extens nombre d’espècies, està demostrant que té un potencial enorme.
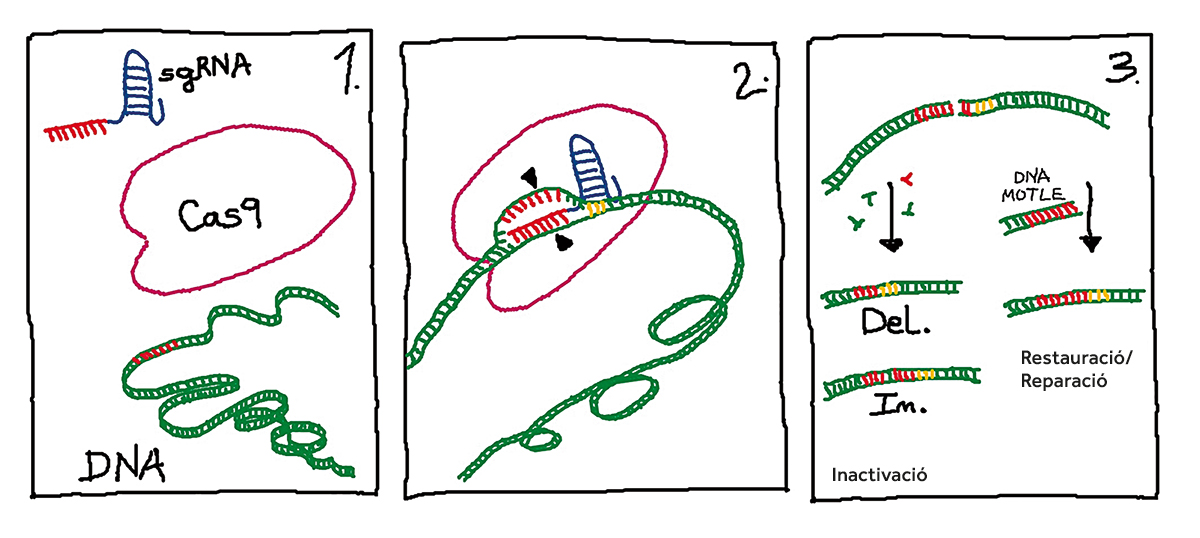
Figura 2. Etapes bàsiques de l’edició de gens mitjançant la tecnologia CRISPR/Cas9. En l’etapa 1, els elements d’edició (sgRNA i proteïna Cas9) s’introdueixen en les cèl·lules a editar on es troba el material genòmic (DNA). El sgRNA porta una seqüència de 20 nucleòtids o seqüència guia (en color roig) complementària al gen diana. En l’etapa 2, el complex sgRNA/Cas9 interacciona amb el DNA genòmic i es produeix l’aparellament entre la seqüència guia del sgRNA i el gen diana. La proteïna Cas9 reconeix una seqüència de tres nucleòtids denominada PAM (en color groc) i realitza un tall en la doble cadena de DNA. En l’etapa 3, la maquinària cel·lular activa els mecanismes cel·lulars de reparació. En el costat esquerre la reparació del tall pot produir-se a costa d’introduir-se (In.: insercions) o perdre’s (Del.: supressions) nucleòtids, la qual cosa generalment porta a la inactivació del gen. Al costat dret es mostra el mecanisme de reparació per recombinació homòloga en el qual es requereix un DNA motle que permet restaurar la seqüència original o introduir un canvi de la nostra elecció. / Concha Gómez Mena
El descobriment dels sistemes CRISPR/Cas es remunta als estudis de sistemes d’immunitat adaptativa microbiana (Mojica i Montoliu, 2016). Alguns microorganismes, principalment arqueus, es protegeixen davant dels virus tallant el seu material genètic. Fragments d’aquest material genètic extern es mantenen en el genoma bacterià com un record de la invasió i, en cas d’una nova infecció, guiaran les nucleases (proteïnes Cas) per al seu tall i inactivació. Eric Lander (2016) va publicar un resum detallat dels principals investigadors implicats en el descobriment i estudi del mecanisme molecular d’aquest sistema.
«La tecnologia CRISPR/Cas fa possible la millora de varietats vegetals sense la introducció d’elements forans»
El desenvolupament de la tecnologia CRISPR/Cas9 per a l’edició de gens en eucariotes es basa en la reconstitució in vitro de tres dels elements identificats en Streptococcus pyogenes, dos RNA (crRNA i tracrRNA) i la proteïna Cas9. Els crRNA (CRISPR RNA) contenen les seqüències homòlogues a gens diana presents en el genoma a editar, mentre que els tracrRNA serveixen d’ancoratge a la proteïna Cas9. La proteïna Cas9 és una endonucleasa que ocasiona talls en la doble cadena de DNA. L’únic requisit per al tall és la presència d’una seqüència de tres nucleòtids denominada PAM3 en la regió adjacent a la seqüència diana (Mojica, Díez-Villaseñor, García-Martínez i Almendros, 2009). El tall de la doble cadena de DNA és reparat per la maquinària cel·lular mitjançant l’eliminació i introducció aleatòria de nucleòtids que generen llocs d’aparellament nous entre les dues cadenes. Si la reparació es produeix a l’entorn de la seqüència codificant d’un gen, els canvis generalment comporten la inactivació d’aquest (Figura 2). Un segon mecanisme permet la reparació del tall a partir d’un DNA motle en un procés de recombinació homòloga (Figura 2). Aquesta segona via fa possible la modificació de seqüències específiques que podria tenir aplicacions en les ciències biomèdiques com a estratègia de teràpia gènica per al tractament de malalties congènites (Montoliu, 2019).
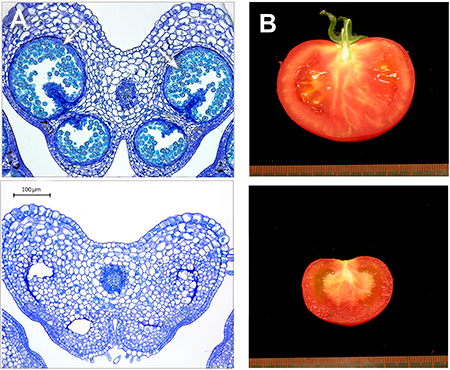
Figura 3. Obtenció de plantes de tomaca sense pol·len mitjançant la tecnologia CRISPR/Cas9. La planta editada (a baix) presenta la inserció d’un nucleòtid en la seqüència d’un gen de tomaca necessari en el procés de gametogènesi masculina. A) Talls histològics d’anteres de flors de tomaca. En les plantes convencionals (a dalt) s’observa la presència del pol·len (fletxes), mentre que les plantes editades (a baix) són estèrils i no produeixen pol·len. B) Fruits madurs d’una tomaca de la varietat Moneymaker convencional (a dalt) i d’una planta editada (a baix). En les plantes editades s’obtenen fruits sense llavors (partenocàrpics). / Fotos de Mª Jesús López Martín
En 2012 el treball de dos laboratoris va proposar la fusió dels dos RNA en una única molècula denominada sgRNA (single-guide RNA) i va demostrar que és funcional i dirigeix la proteïna Cas9 a la zona del genoma on es troba el lloc a editar (Gasiunas, Barrangou, Horvath i Siksnys, 2012; Jinek et al., 2012). Els autors també van predir el potencial d’aquest sistema com a eina per a l’edició genètica programable mediat per nucleases (Jinek et al., 2012). No passaria molt de temps fins a l’aparició de les primeres publicacions d’edició de gens d’organismes in vivo. En 2013 es publiquen els primers articles en què es descriu l’edició de gens en cèl·lules de mamífers (Cong et al., 2013) i plantes (Shan et al., 2013).
Des de llavors la utilització d’aquesta tecnologia en l’edició de gens de diferents organismes ha tingut un increment exponencial. Diversos estudis han demostrat que, utilitzant una sola nucleasa i diversos sgRNA, es poden editar diversos llocs del genoma simultàniament, la qual cosa és especialment interessant quan es pretén editar gens molt grans o famílies de gens (Wolter, Schindele i Puchta, 2019). L’edició de gens en plantes sovint requereix una etapa inicial de transformació genètica per a introduir els elements del sistema (sgRNA i proteïna Cas9) i la posterior eliminació d’aquests en la següent generació. Això significa una limitació per a espècies en què no s’han desenvolupat protocols eficients de transformació genètica o amb llargs temps de generació, com algunes plantes llenyoses. En aquest sentit s’estan desenvolupat mètodes alternatius que no requereixen transformació estable utilitzant protoplasts o embrions immadurs (Liang et al., 2017; Osakabe et al., 2018). També s’han desenvolupat sistemes d’edició compostos únicament per proteïnes o complexos RNA-proteïna que permeten editar el genoma sense l’ús de DNA recombinant (Metje-Sprink, Menz, Modrzejewski i Sprink, 2018).
El futur dels aliments i l’edició genètica en plantes
En plantes de cultiu, el sistema CRISPR/Cas9 s’ha utilitzat en la millora de cereals com ara dacsa, arròs i blat; hortícoles –tomaca, creïlla, cogombre i meló d’Alger– i espècies llenyoses –cítrics, papaia i pomera, entre d’altres– (revisat per Wang, Zhang i Zhu, 2019; Wolter et al., 2019).
La tecnologia CRISPR/Cas9 s’ha usat extensament per a generar plantes resistents a estressos tant abiòtics (condicions ambientals adverses que afecten el creixement de la planta) com biòtics (microorganismes patògens). Recentment, un grup del Centre Nacional de Biotecnologia a Madrid ha reportat que l’edició del gen SlJAZ2 de tomaca, relacionat amb la percepció de l’hormona vegetal jasmonat, confereix protecció a la planta davant de fongs patògens per mitjà d’un mecanisme associat al tancament estomàtic (Ortigosa, Giménez-Ibañez, Leonhardt i Solano, 2019). En cereals com l’arròs, gran part de les millores introduïdes mitjançant la tecnologia CRISPR/Cas9 s’han concentrat en el desenvolupament de plantes més resistents a fongs fitopatògens i a incrementar la resistència a altes temperatures (Biswal, Mangrauthia, Reddy i Yugandhar, 2019) associades a canvi climàtic.
«L’edició genètica a la carta té un enorme potencial com a nova estratègia en la millora dels cultius del futur»
Un exemple d’edició genètica amb important repercussió en la salut humana és el desenvolupament de plantes de blat amb baix contingut en proteïnes del gluten, responsables de diferents trastorns del sistema digestiu (Sánchez-León et al., 2018). En aquest treball els autors dissenyen dos sgRNA per a editar regions conservades dels gens que codifiquen les alfa–gliadines, les principals proteïnes que desencadenen sensibilitat en persones que pateixen de malaltia celíaca. L’aproximació va reeixir en la introducció de mutacions de manera específica i simultània en aquesta família de proteïnes, i es van obtenir plantes amb baix contingut en gluten i una disminució en la seua immunoreactivitat del 85 % (Sánchez-León et al., 2018). Les línies obtingudes constitueixen un avanç sense precedents en l’obtenció de varietats comercials de blat amb baix contingut en gluten en un futur pròxim.
Al nostre laboratori de l’Institut de Biologia Molecular i Cel·lular de Plantes (IBMCP, València) hem estudiat durant els últims anys el procés de formació del fruit en plantes de tomaca. Els nostres estudis revelen un mecanisme pel qual les anteres en desenvolupament regulen negativament el creixement de l’ovari no pol·linitzat. En conseqüència, l’eliminació primerenca dels estams té com a conseqüència el desenvolupament de fruits sense llavors en aquesta espècie (Medina et al., 2013; Rojas-Gracia et al., 2017).
Basant-nos en aquests resultats, hem utilitzat la tecnologia CRISPR/Cas9 per a demostrar que la inactivació específica de gens responsables de la formació del pol·len és suficient per a desencadenar el desenvolupament de fruits sense llavors (partenocàrpics). En concret, la inserció d’un únic nucleòtid en la seqüència d’un gen diana inactiva la seua funció del gen i dona lloc a plantes sense pol·len i partenocàrpiques (Figura 3; resultats sense publicar). Aquest desenvolupament podria tenir altres aplicacions en l’obtenció de plantes ornamentals sense pol·len com a mètode de contenció per a evitar-ne la propagació indesitjada o per a reduir-hi la presència d’al·lergògens. Aquestes noves varietats són un exemple d’edició genètica a la carta i demostren l’enorme potencial d’aquestes noves estratègies en la millora dels cultius del futur.
Notes
1. Vegeu el web http://www.theplantlist.org/ (Torna al text)
2. Les sigles CRISPR/Cas (CRISPER-associated) signifiquen clustered regularly interspaced short palindromic repeats (“repeticions palindròmiques curtes agrupades i regularment espaiades”). (Torna al text)
3. Protospacer adjacent motif o “motiu adjacent de protoespaciador”. En el cas de S. pyogenes s’ha determinat que la seqüència PAM és NGG, on N és qualsevol nucleòtid i G un nucleòtid de guanina. (Torna al text)
Referències
Beltrán, J. P. (2018). Cultivos transgénicos. Madrid: CSIC-Los libros de la Catarata.
Biswal, A. K., Mangrauthia, S. K., Reddy, M. R., & Yugandhar, P. (2019). CRISPR mediated genome engineering to develop climate smart rice: Challenges and opportunities. Seminars in Cell & Developmental Biology, 96, 100–106. doi: 10.1016/j.semcdb.2019.04.005
Cong, L., Ran, F. A., Cox, D., Lin, S., Barretto, R., Habib, N., . . . Zhang, F. (2013). Multiplex genome engineering using CRISPR/Cas systems. Science, 339(6121), 819–823. doi: 10.1126/science.1231143
FAO. (1983). World food security: A reappraisal of the concepts and approaches. Roma: Food and Agriculture Organization of the United Nations.
FAO. (1999). Women: Users, preservers and managers of agrobiodiversity. Roma: Food and Agriculture Organization of the United Nations.
FAO, FIDA, & PMA. (2012). El estado de la inseguridad alimentaria en el mundo. Roma: Organización de las Naciones Unidas para la Alimentación y la Agricultura.
Gasiunas, G., Barrangou, R., Horvath, P., & Siksnys, V. (2012). Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proceedings of the National Academy of Science of the USA, 109(39), E2579–2586. doi: 10.1073/pnas.1208507109
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., & Charpentier, E. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science, 337(6096), 816–821. doi: 10.1126/science.1225829
Lander, E. S. (2016). The Heroes of CRISPR. Cell, 164(1-2), 18–28. doi: 10.1016/j.cell.2015.12.041
Liang, Z., Chen, K., Li, T., Zhang, Y., Wang, Y., Zhao, Q., . . . Gao, C. (2017). Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nature Communications, 8, 14261. doi: 10.1038/ncomms14261
Medina, M., Roque, E., Pineda, B., Cañas, L., Rodríguez-Concepción, M., Beltrán, J. P., & Gómez-Mena, C. (2013). Early anther ablation triggers parthenocarpic fruit development in tomato. Plant Biotechnology Journal, 11(6), 770–779. doi: 10.1111/pbi.12069
Metje-Sprink, J., Menz, J., Modrzejewski, D., & Sprink, T. (2018). DNA-free genome editing: Past, present and future. Frontiers in Plant Science, 9, 1957. doi: 10.3389/fpls.2018.01957
Mojica, F. J., Díez-Villaseñor, C., García-Martínez, J., & Almendros, C. (2009). Short motif sequences determine the targets of the prokaryotic CRISPR defence system. Microbiology, 155(Pt 3), 733–740. doi: 10.1099/mic.0.023960-0
Mojica, F. J., & Montoliu, L. (2016). On the origin of CRISPR-Cas technology: From prokaryotes to mammals. Trends in Microbiology, 24(10), 811–820. doi: 10.1016/j.tim.2016.06.005
Montoliu, L. (2019). Editando genes: recorta, pega y colorea. Las maravillosas herramientas CRISPR. Pamplona: Next Door Publishers.
National Academies of Sciences & Medicine. (2016). Genetically engineered crops: Experiences and prospects. Washington, DC: The National Academies Press.
Ortigosa, A., Giménez-Ibáñez, S., Leonhardt, N., & Solano, R. (2019). Design of a bacterial speck resistant tomato by CRISPR/Cas9-mediated editing of SlJAZ2. Plant Biotechnology Journal, 17(3), 665–673. doi: 10.1111/pbi.13006
Osakabe, Y., Liang, Z., Ren, C., Nishitani, C., Osakabe, K., Wada, M., . . . Nagamangala Kanchiswamy, C. (2018). CRISPR-Cas9-mediated genome editing in apple and grapevine. Nature Protocols, 13(12), 2844–2863. doi: 10.1038/s41596-018-0067-9
Rojas-Gracia, P., Roque, E., Medina, M., Rochina, M., Hamza, R., Angarita-Díaz, M. P., . . . Gómez-Mena, C. (2017). The parthenocarpic hydra mutant reveals a new function for a SPOROCYTELESS-like gene in the control of fruit set in tomato. New Phytologist, 214(3), 1198–1212. doi: 10.1111/nph.14433
Sánchez-León, S., Gil-Humanes, J., Ozuna, C. V., Giménez, M. J., Sousa, C., Voytas, D. F., & Barro, F. (2018). Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnology Journal, 16(4), 902–910. doi: 10.1111/pbi.12837
Shan, Q., Wang, Y., Li, J., Zhang, Y., Chen, K., Liang, Z., . . . Gao, C. (2013). Targeted genome modification of crop plants using a CRISPR-Cas system. Nature Biotechnology, 31(8), 686–688. doi: 10.1038/nbt.2650
Taylor, S. L., & Hefle, S. L. (2001). Ingredient and labeling issues associated with allergenic foods. Allergy: European Journal of Allergy and Clinical Immunology, 56(67), 64–69. doi: 10.1034/j.1398-9995.2001.00920.x
Wang, T., Zhang, H., & Zhu, H. (2019). CRISPR technology is revolutionizing the improvement of tomato and other fruit crops. Horticulture Research, 6(1), 77. doi: 10.1038/s41438-019-0159-x
Wolter, F., Schindele, P., & Puchta, H. (2019). Plant breeding at the speed of light: the power of CRISPR/Cas to generate directed genetic diversity at multiple sites. BMC Plant Biology, 19(1), 176. doi: 10.1186/s12870-019-1775-1




