

Més enllà del recompte d’espècies
Una nova manera d’enfocar la biodiversitat

Els índexs tradicionals de diversitat (principalment riquesa, abundància i equitativitat d’espècies) han estat altament reveladors per al seguiment de processos en comunitats i ecosistemes en ecologia moderna. No obstant això, fa aproximadament dues dècades, un grup d’investigadors, pioners en el seu camp, van apreciar que aquests índexs no resolien per complet les qüestions que es proposaven. Per això van plantejar canviar la manera de quantificar la biodiversitat. Fonamentalment, aquesta nova metodologia considera la distància entre les espècies (en termes filogenètics o funcionals) de manera prèvia a aplicar els índexs de biodiversitat oportuns. La incorporació d’elements filogenètics i funcionals en l’avaluació de la diversitat permet acostar-nos al concepte de biodiversitat de manera més completa.
Paraules clau: índexs clàssics de diversitat, diversitat funcional, diversitat filogenètica, trets funcionals, distàncies genètiques.
La biodiversitat o diversitat biològica és un concepte que fa referència a la varietat de la vida sobre la Terra com a resultat de milers de milions d’anys d’evolució. Probablement la primera imatge que ve a la nostra ment en sentir aquesta paraula són escenaris idealitzats: selves tropicals d’arbres verds, llenyosos i frondosos, poblades per mamífers i aus exòtiques, i potser inclourem en la nostra foto mental algun insecte vistós, com una papallona de colors cridaners. O, tal vegada, evocarem un escull de coral en què naden peixos combinats en colors impossibles. No obstant això, la dimensió «evolutiva» del terme biodiversitat ens fa sospitar que deu haver-hi alguna cosa més en aquest escenari.
«Per a gestionar la biodiversitat és fonamental realitzar una quantificació fiable que tinga en compte els diferents nivells d’organització»
La vida sobre la Terra comprèn un gran nombre de grups vegetals i animals, però també molts més eucariotes (organismes amb cèl·lules nucleades) i encara més grups de procariotes (organismes amb cèl·lules sense nucli). Totes aquestes formes de vida són protagonistes dels processos que es donen en els ecosistemes. És per aquests processos pels quals conèixer la biodiversitat del medi natural ha estat una inquietud constant per a l’ésser humà. Primer, per a posar-la al seu servei o extraure els seus recursos; després, per a conservar-la per raons que poden anar més enllà del mer utilitarisme, incorporant una dimensió eudemònica a la conservació, i, durant les últimes dècades, per a avaluar i mitigar l’impacte de les pertorbacions lligades al canvi climàtic sobre la vida en el planeta. La biodiversitat engloba variabilitat a tres nivells: «entre ecosistemes»; entre les unitats taxonòmiques (per simplificar, d’ara en avant espècies)1 que habiten en els ecosistemes («interespecífica»), i dins de cadascuna de les espècies («intraespecífica») (Glowka, Burhenne-Guilmin, Synge, McNeely i Gündling, 1994). Per això, un dels pilars fonamentals per a gestionar-la és realitzar una quantificació fiable que tinga en compte aquests nivells d’organització.
Descomposició espaciotemporal de la biodiversitat
Històricament, els ecòlegs prompte van ser conscients que la variabilitat biològica pot presentar patrons diferents depenent de l’escala considerada. Whittaker (1960) va ser el primer a descriure els diferents components espacials en què es pot mesurar la biodiversitat. Aquest autor va proposar abordar l’estudi de la biodiversitat al llarg d’una sèrie d’escales espacials jeràrquiques (Figura 1). Així va definir la diversitat gamma (γ) com la diversitat d’espècies en una regió o ecosistema. Per la seua banda, l’escala jeràrquica més baixa, corresponent al punt de mostreig, va rebre el nom de la diversitat alfa (α). Finalment, va definir la diversitat beta (β), que estableix la dissemblança entre dos mòduls comparables, normalment corresponent a un mateix nivell jeràrquic, com per exemple la dissemblança de biodiversitat entre diversos punts de mostreig.
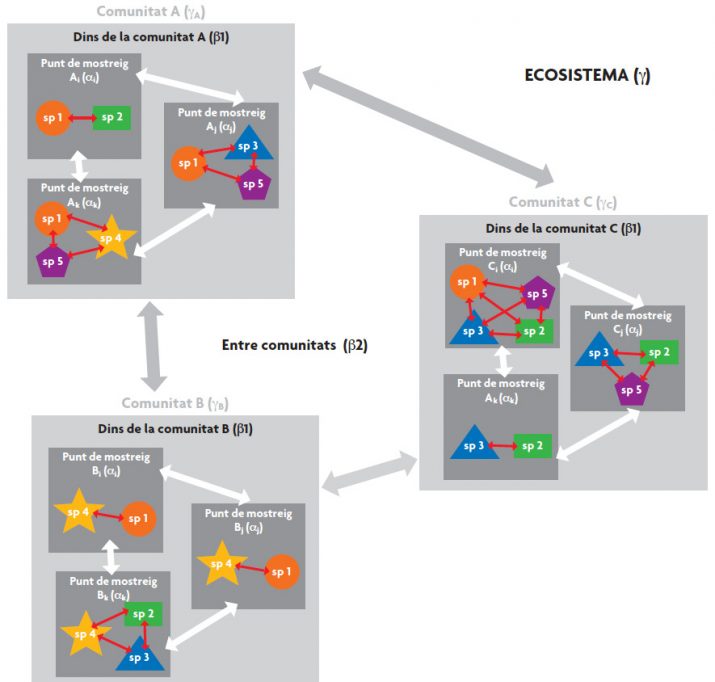
A causa de la seua gran utilitat i a l’extensió d’ús, posteriorment va sorgir la necessitat de redefinir el component beta. Així doncs, es va fer la distinció entre diversitat beta 1 (β1), com la dissemblança entre els punts de mostreig dins d’una mateixa comunitat; i diversitat beta 2 (β2), com la dissemblança entre les comunitats d’una regió (Excoffier, Smouse i Quattro, 1992) (vegeu Figura 1). Actualment, els últims estudis en aquest camp proposen una descomposició temporal de la biodiversitat, anàloga a l’espacial. D’aquesta manera, l’escala jeràrquica es podria estendre a mòduls temporals niats, com per exemple anys, dècades i segles. Una conseqüència lògica d’aquest disseny és la possibilitat de combinar els aspectes espaciotemporals de la biodiversitat, però aquest enfocament està encara poc desenvolupat (Pavoine i Bonsall, 2011).
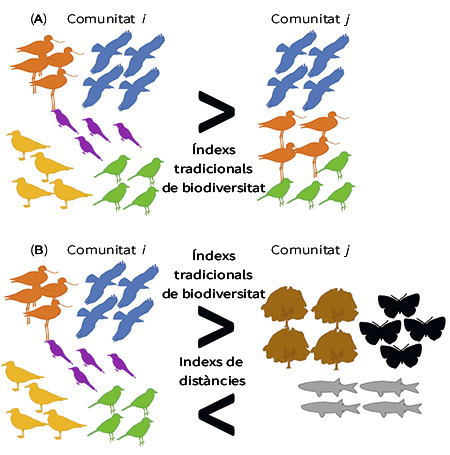
Figura 2. Les figures A i B mostren dues comunitats amb una mateixa abundància en totes les espècies presents, encara que amb una major riquesa d’espècies en la comunitat i que en la comunitat j en ambdós casos. No obstant això, en la comunitat j de la figura B s’han reemplaçat les espècies per altres filogenèticament o funcionalment distants. D’acord amb els índexs tradicionals de biodiversitat, tant en (A) com en (B) la comunitat i és més diversa que la comunitat j. Mentre que, d’acord amb els índexs de distàncies (diversitat filogenètica i diversitat funcional), en (B) la comunitat j és més diversa que la comunitat i.
Com mesurem la biodiversitat?
Independentment de l’escala espaciotemporal (α, β, γ), els índexs que s’han utilitzat per a quantificar i caracteritzar la diversitat s’han basat primordialment a avaluar la variabilitat a nivell «interespecífic», per ser aquest molt més fàcil d’apreciar i quantificar. En canvi, les diversitats «intraespecífica» i d’«ecosistemes» han rebut molta menor atenció. Els índexs tradicionals de biodiversitat quantifiquen principalment la riquesa, l’abundància i l’equitativitat de les espècies d’interès presents en una mostra. Així, donada una mostra definida a escala espacial o temporal, la riquesa d’espècies indica el nombre d’espècies presents, l’abundància quantifica el nombre d’individus de cada espècie i, finalment, l’equitativitat relaciona la riquesa i l’abundància, de manera que estableix el grau en què els individus es reparteixen entre les espècies comptades en la mostra. Gràcies a aquests índexs tradicionals s’ha pogut caracteritzar la biodiversitat de manera fàcil i intuïtiva a qualsevol escala espacial (α, γ) i fins i tot realitzar comparacions entre mòduls del mateix nivell jeràrquic (β).
Tanmateix, aquests índexs no ofereixen tots els matisos necessaris per a obtenir una idea completa de la biodiversitat, perquè desatenen dues de les seues dimensions: la filogenètica i la funcional. Per exemple, si apliquem els índexs tradicionals a dues comunitats (comunitat i i comunitat j, tal com observem en la Figura 2a), en les quals trobem la mateixa abundància en totes les espècies presents i amb una riquesa d’espècies Si = 5 i Sj = 3, l’equitativitat serà Ei = 100 % i Ej = 100 %.
No obstant això, quan reemplacem les espècies de la comunitat j per d’altres de filogenèticament (genèticament) o funcionalment (que aporten i ocupen serveis molt diferents en l’ecosistema) distants (Figura 2b), els índexs tradicionals de biodiversitat continuaran proporcionant-nos els mateixos resultats relatius. És a dir, la comunitat i serà més diversa que la j, sense considerar com són de diferents les espècies de les comunitats. Aquest exemple clarament mostra una limitació fonamental dels índexs de biodiversitat tradicionals. Per aquesta raó, durant les últimes dècades, un bon nombre d’ecòlegs s’han esforçat per definir un nou marc matemàtic que descriga les diferències filogenètiques i funcionals de les espècies d’una comunitat.
Entre les primeres aproximacions, Faith (1992) va intentar avaluar i considerar les distàncies filogenètiques entre espècies. Aquest autor va exposar que, abans de mesurar la biodiversitat d’una mostra donada, s’haurien de calibrar les distàncies filogenètiques de les espècies que componen la mostra. D’aquesta manera, es podria conèixer la singularitat evolutiva de cada llinatge (separació més o menys pròxima en el temps). Així, va proposar mesurar el parentiu filogenètic entre les espècies d’una mostra com una matriu entre parells d’espècies. Aquesta matriu s’extrau de la distància en un arbre filogenètic entre cada parell d’espècies. D’aquesta manera, diem que qualsevol estudi que base les seues mesures de biodiversitat en les distàncies filogenètiques entre espècies serà un estudi de diversitat filogenètica.
«Els ecòlegs prompte van ser conscients que la variabilitat biològica pot presentar patrons diferents depenent de l’escala considerada»
Posteriorment Petchey i Gaston (2002) van plantejar un protocol anàleg al de Faith (1992), però en aquest cas les relacions entre espècies no es construeixen a partir de seqüències filogenètiques, sinó a partir de trets funcionals.2 Els trets funcionals són les unitats utilitzades per a mesurar la diversitat funcional d’un conjunt d’espècies i permeten avaluar les conseqüències d’una gran varietat de qüestions ecològiques, per exemple: l’impacte del canvi climàtic en la diversitat o la successió ecològica després de la restauració d’un hàbitat. En aquest cas, es mesuraria la complementarietat funcional entre les espècies de la nostra mostra, per a construir una matriu de distàncies entre cada parell d’espècies, a partir de dades de trets funcionals. En creuar aquesta matriu amb l’índex adequat de biodiversitat, sabrem com és de diversa la nostra mostra en termes de diversitat funcional.
Si finalment tornem a examinar la Figura 2b i, prèviament a mesurar la biodiversitat de cada mostra, calibrem la distància entre les espècies de les nostres comunitats en termes de distàncies filogenètiques (diversitat filogenètica) o funcionals (diversitat funcional), veurem que la nostra conclusió sobre quina és la comunitat més diversa es veu alterada per complet.
Què han aportat aquests estudis?
Dels estudis de les últimes dècades, podem extraure afirmacions tan cridaneres com ara que un aparatós escull de coral és menys biodivers que un auster ecosistema de muntanya (Figura 3). Si bé en el primer s’ha vist afavorida la radiació o aparició de noves espècies, aquestes són filogenèticament molt pròximes i pràcticament iguals a escala funcional. Per això, si afegim o extraiem una espècie de l’ecosistema, els valors de diversitat filogenètica o funcional romandran pràcticament immutables. Per contra, l’ecosistema de muntanya té un menor nombre d’espècies, però aquestes són filogenèticament i funcionalment molt distants, per la qual cosa perdre una de les espècies comportaria una caiguda dràstica dels valors de diversitat filogenètica o funcional.

Figura 3. D’acord amb els índexs tradicionals de diversitat, la comunitat de peixos d’un escull de coral (dreta) seria més biodiversa que tot un ecosistema de muntanya (esquerra), perquè el primer té un nombre més gran d’espècies. No obstant això, aquestes espècies són filogenèticament (evolutivament) i funcionalment (ecològicament) semblants, per la qual cosa són redundants en termes de diversitat filogenètica i funcional. Mentre que en l’ecosistema de muntanya, fins i tot amb un menor nombre d’espècies, cadascuna d’elles és filogenèticament i funcionalment diferent de la resta. / Adi Ulici, NOAA
No pretenem indicar amb açò que un ecosistema coral·lí no siga igual de mereixedor de ser conservat que un d’alta muntanya. El que volem transmetre, però, és que la deguda consideració dels aspectes funcionals i filogenètics de les comunitats biològiques ens ajuda a entendre millor l’organització de la biodiversitat en el planeta i s’ha de tenir en compte a l’hora d’establir mesures concretes de conservació.
Pròxima parada?
La realització d’aquests estudis és més complexa que en el cas de les aproximacions tradicionals, ja que requereixen afrontar dos reptes en primera instància. El primer és la identificació de trets funcionals significatius i no redundants per a quantificar la diversitat funcional. El segon és la disponibilitat d’informació sobre les relacions de parentiu entre les espècies per a poder quantificar la diversitat filogenètica. No obstant això, és interessant assenyalar que, a diferència dels índexs de biodiversitat tradicionals, els conceptes de diversitat filogenètica i funcional són directament aplicables a escala d’individu. A pesar d’haver estat poc explorada, aquesta aproximació possibilita l’extensió a estudis de diversitat intraespecífica, la qual cosa permet la integració d’aquest aspecte en estudis de biodiversitat. Per exemple, mesurar la diversitat filogenètica intraespecífica podria ser crucial a l’hora d’entendre els patrons filogeogràfics i reconeixement de subespècies, per a així implantar plans de conservació biològica (Excoffier, 2008).
«Dels estudis de les últimes dècades, podem extraure afirmacions tan cridaneres com ara que un aparatós escull de coral és menys biodivers que un auster ecosistema de muntanya»
A més, els índexs de diversitat filogenètica i funcional han servit per a caracteritzar un gran nombre d’ecosistemes terrestres i aquàtics, prenent com a model des de vegetals herbacis o llenyosos a insectes i vertebrats. No obstant això, considerem que encara queden molts camps d’aplicació. Per exemple, els paràsits, per la seua forma de vida que depén d’un altre organisme, tenen característiques que els fan idonis per a revelar processos ocults en els ecosistemes. Així mateix, els paràsits són ubics en tots els ecosistemes; alguns tenen cicles de vida complexos, per la qual cosa són útils per a traçar rutes en les xarxes tròfiques i dilucidar patrons espaciotemporals dels ecosistemes (Poulin i Morand, 2000). A pesar d’això, pocs autors han abordat l’estudi de la biodiversitat de les comunitats de paràsits des del punt de vista filogenètic o funcional. Els estudis més recents indiquen que els organismes paràsits realitzen funcions reguladores, protectores i estabilitzadores en els ecosistemes. A més, l’estudi dels sistemes paràsit-hoste aporta un potent instrument comparatiu, a causa de l’organització niada3 de les comunitats paràsites, que pot proporcionar conclusions generalitzables a altres comunitats biològiques.
Conclusió
Actualment, és difícil concebre un estudi que explique o prediga els processos que tenen lloc en els ecosistemes sense incloure dades filogenètiques o funcionals de les unitats taxonòmiques considerades en la mostra. No obstant això, ens agradaria destacar que els estudis basats en la comparació dels resultats obtinguts mitjançant diferents índexs de biodiversitat, així com a diferents escales jeràrquiques d’organització, poden revelar processos evolutius, biogeogràfics o de radiació que altrament passarien inadvertits. Per això, convidem els investigadors interessats a utilitzar aquest nou marc conceptual en els seus estudis.
1. En el text simplifiquem la descripció d’unitat taxonòmica equiparant-la a la descripció d’espècie biològica. No obstant això, cal destacar que els índexs de biodiversitat, tant els tradicionals com els basats en distàncies filogenètiques o funcionals, poden aplicar-se a qualsevol nivell de la classificació taxonòmica dels organismes i fins i tot a virus (Shi et al., 2016), que en certs àmbits s’escapen de la definició d’organisme biològic. (Tornar)
2. D’acord amb Carmona, De Bello, Mason & Lepš (2016), un tret funcional és qualsevol característica morfològica, fisiològica, fenològica o comportamental, que es pot mesurar a escala d’individu i té un efecte sobre la seua supervivència o reproducció. (Tornar)
3. Les anàlisis de conjunts niats (per exemple, associacions paràsit-hoste) poden revelar patrons ecològics no aleatoris i són eines exploradores útils per a suggerir mecanismes que poden estructurar una comunitat en particular (González i Poulin, 2005). (Tornar)
Referències
Carmona, C. P., De Bello, F., Mason, N. W. H., & Lepš, J. (2016). Traits without borders: Integrating functional diversity across scales. Trends in Ecology & Evolution, 31(5), 382–394. doi: 10.1016/j.tree.2016.02.003
Excoffier, L. (2008). Analysis of population subdivision. In D. J. Balding, M. Bishop, & C. Cannings (Eds.), Handbook of statistical genetics (pp. 980–1020). Chichester: John Wiley & Sons Ltd. doi: 10.1002/9780470061619.ch29
Excoffier, L., Smouse, P. E., & Quattro, J. M. (1992). Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics, 131(2), 479–491.
Faith, D. P. (1992). Conservation evaluation and phylogenetic diversity. Biological Conservation, 61(1), 1–10. doi: 10.1016/0006-3207(92)91201-3
Glowka, L., Burhenne-Guilmin, F., Synge, H., McNeely, J. A., & Gündling, L. (1994). A guide to the convention on biological diversity (N.º 333.95 G946). Gland/Cambdrige/Bonn: IUCN and Environmental Law Centre IUCN.
González, M. T., & Poulin, R. (2005). Nested patterns in parasite component communities of a marine fish along its latitudinal range on the Pacific coast of South America. Parasitology, 131(4), 569–577. doi: 10.1017/S0031182005007900
Pavoine, S., & Bonsall, M. B. (2011). Measuring biodiversity to explain community assembly: A unified approach. Biological Reviews, 86(4), 792–812. doi: 10.1111/j.1469-185X.2010.00171.x
Petchey, O. L., & Gaston, K. J. (2002). Functional diversity (FD), species richness and community composition. Ecology Letters, 5(3), 402–411. doi: 10.1046/j.1461-0248.2002.00339.x
Poulin, R., & Morand, S. (2000). The diversity of parasites. The Quarterly Review of Biology, 75(3), 277–293. doi: 10.1086/393500
Shi, M., Lin, X. D., Tian, J. H., Chen, L. J., Chen, X., Li, C. X., ... Zhang, Y.-Z. (2016). Redefining the invertebrate RNA virosphere. Nature, 540(7634), 539–543. doi: 10.1038/nature20167
Whittaker, R. H. (1960). Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monographs, 30(3), 279–338. doi: 10.2307/1943563




